Article Text

Abstract
Objective Zinc-finger protein 545 (ZNF545) is a member of the family of Krüppel-associated box-containing zinc-finger proteins. The aim of this study was to clarify its biological function as a tumour suppressor in gastric cancer.
Design The biological function of ZNF545 was determined by cell growth and apoptosis assays. The ZNF545 target signal pathway was identified by promoter luciferase assay, northern blot, run-on transcription assay, chromatin immunoprecipitation and coimmunoprecipitation assays. The clinical application of ZNF545 was assessed in primary gastric cancers.
Results ZNF545 was silenced or reduced in 16 out of 18 gastric cancer cell lines by promoter hypermethylation. Restoration of ZNF545 expression in gastric cancer cell lines suppressed cell proliferation and induced apoptosis. These effects of ZNF545 were attributed to inhibition of ribosomal RNA (rRNA) transcription. Inhibition of rRNA transcription by ZNF545 was further revealed to be associated with direct ribosomal DNA (rDNA) promoter binding, recruitment of the corepressor, heterochromatin protein 1β, and reduction of trimethylated histone H3 at the Lys4 residue at the rDNA locus. ZNF545 methylation was detected in 51.9% (41/79) of gastric cancer tissues, 27.0% (20/74) of adjacent non-tumour gastric tissues (p=0.001), but none of 20 normal controls. Multivariate analysis revealed that patients with ZNF545 methylation had a significant decrease in overall survival. Kaplan–Meier survival curves showed that ZNF545 methylation was significantly associated with shortened survival in patients with stage I–II gastric cancer.
Conclusions ZNF545 acts as a functional tumour suppressor in gastric cancer by inhibiting rRNA transcription. Its methylation at early stages of gastric carcinogenesis is an independent prognostic factor.
- Zinc finger protein 545
- tumour suppressor
- rRNA transcription
- gastric cancer
- prognosis
- cancer
- tumour markers
- methylation
- gene regulation
- molecular carcinogenesis
- molecular oncology
- colorectal adenomas
- gastrointestinal bleeding
- colorectal cancer screening
- non-alcoholic steatohepatitis
Statistics from Altmetric.com
- Zinc finger protein 545
- tumour suppressor
- rRNA transcription
- gastric cancer
- prognosis
- cancer
- tumour markers
- methylation
- gene regulation
- molecular carcinogenesis
- molecular oncology
- colorectal adenomas
- gastrointestinal bleeding
- colorectal cancer screening
- non-alcoholic steatohepatitis
Significance of this study
What is already known on this subject?
-
Epigenetic alterations, particularly inactivation of tumour-suppressor genes by promoter hypermethylation, play an important role in the initiation and progression of gastric cancer.
-
The ZNF545 gene, located on chromosome 19q13.12, produces a protein that is a member of the family of Krüppel-associated box-containing zinc-finger proteins, which may act in the nucleus as a sequence-specific transcriptional repressor.
-
Expression of human ZNF545 mRNA is ubiquitously detected in human tissues.
What are the new findings?
-
ZNF545 is commonly silenced or downregulated in gastric cancer by promoter methylation.
-
ZNF545 acts as a novel tumour suppressor in gastric cancer.
-
The anti-tumorigenic function of ZNF545 was modulated by inhibiting rRNA transcription by direct binding to rDNA promoter, inducing histone modification changes, and recruiting the corepressor, heterochromatin protein 1β.
-
ZNF545 methylation is a potential epigenetic biomarker for predicting outcome of patients with gastric cancer.
How might it impact on clinical practice in the foreseeable future?
-
Detection of methylated ZNF545 may serve as a new biomarker for the prognosis of patients with gastric cancer.
Introduction
Gastric cancer is one of the most common malignancies and remains the second leading cause of cancer-related death worldwide. Although the mechanism leading to gastric cancer development remains elusive, epigenetic inactivation of tumour-related genes by promoter hypermethylation has been demonstrated to play an important role in the development of this malignancy. The identification of novel genes targeted by promoter methylation may help us to unravel the molecular mechanisms involved in inactivation of tumour suppressive pathways and pathogenesis of gastric cancer and to find better approaches to diagnostic and therapeutic evaluation.
Krüppel-associated box domain-zinc-finger proteins (KRAB-ZFPs) constitute a large subfamily of zinc-finger proteins, which has become the largest family of transcriptional regulators in the human genome.1 KRAB-ZFP family members have been found to mediate transcriptional repression of RNA polymerases and binding and splicing of ribosomal RNAs (rRNAs).1 They can also mediate transcription silencing by binding to the promoter of target genes through a C2H2 zinc-finger domain and recruiting corepressors such as heterochromatin protein 1 (HP1) through the KRAB domain.1 ,2 Aberrant inactivation of KRAB-ZFPs has been reported to contribute to irregular gene expression and tumour development.3 Characterisation of the functional significance of KRAB-ZFP family members in gastric cancer may provide new insights for understanding the molecular mechanisms of gastric carcinogenesis and identifying potential targets for the diagnosis and treatment of gastric cancer. Through genome-wide screening, we identified a novel KRAB-ZFP member, zinc-finger protein 545 (ZNF545), which is methylated in gastric cancer. However, the effects of ZNF545 in tumorigenicity remain unexplored. In this study, we therefore evaluated the expression profile, epigenetic regulation, biological function, molecular basis and clinical application of ZNF545 in gastric cancer.
Materials and methods
Cell lines
Eighteen gastric cancer cell lines (AGS, Kato III, MKN28, MKN45, N87, SNU1, SNU16, YCC1, YCC2, YCC3, YCC6, YCC7, YCC9, YCC10, YCC11, YCC16, BGC823 and MGC803) and one normal gastric epithelial cell line (GES1) were used in this study. Cell lines were maintained in RPMI 1640 or Dulbecco's modified Eagle's medium (Gibco BRL, Rockville, Maryland, USA) with 10% fetal bovine serum. Human normal adult tissue RNA samples were purchased commercially (Stratagene, La Jolla, California, USA).
Gastric tissue samples
Paired gastric biopsy samples from primary tumour and adjacent non-tumour sites were obtained from patients with gastric cancer during endoscopy before any therapeutic intervention as described previously.4 The adjacent non-tumour area was subsequently verified by histology to be free of tumour infiltration. A total of 79 patients with confirmed gastric cancer and adjacent non-tumour tissues were examined for ZNF545 methylation. Adjacent non-tumour tissue was examined for diagnosis of chronic gastritis, glandular atrophy, intestinal metaplasia, and dysplasia of mucosa as stipulated by the updated Sydney system. The adjacent non-tumour tissues contained 45 samples with chronic gastritis and 34 samples with precancerous lesions (four with atrophy, 28 with intestinal metaplasia, and two with dysplasia). In addition, 20 age-matched subjects with normal upper gastroscopy were recruited as controls. The study protocol was approved by the Clinical Research Ethics Committee of the Chinese University of Hong Kong.
Assay of colony formation
Gastric cancer cells (AGS and MGC803) were transfected with pcDNA3.1-Flag-ZNF545 or empty pcDNA3.1. After 48 h of transfection, cells were selected with G418 (Calbiochem, Darmstadt, Germany) for 10–14 days. Colonies (≥50 cells/colony) were counted after being fixed with 70% ethanol and stained with 5% crystal violet.
Assay of cell viability
Cell viability was determined by the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium assay (Promega, Madison, Wisconsin, USA).
DDAO-SE labelling assay
CellTrace Far Red DDAO-SE dye (Invitrogen, Carlsbad, California, USA) was used as a live cell labelling dye to analyse cell proliferation. Equal numbers of stable ZNF545-expressing cells and control cells were seeded on a six-well plate. After 24 h, cells were incubated with 5 μM DDAO-SE for 10 min at 37°C. Staining was quenched by placing the cells in fresh culture medium and incubating for an additional 30 min at 37°C. Cells were then harvested at the starting time point and at 48 h, and the fluorescence intensity of the DDAO-SE signal, which corresponded to the proliferation status of the cultured cells, was measured by flow cytometry (Becton Dickinson Biosciences, Bedford, Massachusetts, USA).
Assay of apoptosis
Apoptosis was determined by dual staining with annexin V-APC (Becton Dickinson Biosciences) and 7-aminoactinomycin (7-AAD) (Becton Dickinson Biosciences). Stable ZNF545-expressing or control cells (AGS and MGC803) were harvested, stained and analysed by flow cytometry according to the manufacturer's instructions (Becton Dickinson Biosciences). Sample fluorescence of 10 000 cells was analysed using the FACSCalibur System (Becton Dickinson Pharmingen, San Jose, California, USA). The relative proportion of annexin V-positive and 7-AAD-negative cells was determined using ModFitLT software (Becton Dickinson Pharmingen) and counted as early apoptotic cells. All the experiments were performed in triplicate independently.
Fluorescent immunohistochemistry
AGS or MGC803 cells were seeded on coverslips on a six-well plate and transfected with pcDNA3.1-Flag-ZNF545. At 48 h after transfection, cells were fixed with 4% paraformaldehyde and permeabilised with 0.3% Triton X-100, blocked in 3% bovine serum albumin, and stained with monoclonal antibody to Flag M2 (1:50 dilution, F1804; Sigma, St Louis, Missouri, USA) overnight at 4°C. After incubation with Texas Red-conjugated goat anti-rabbit IgG (1:500 dilution, Santa Cruz Biotechnology, Santa Cruz, California, USA), cells were mounted with mounting medium containing 4',6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, California, USA). Images were captured by fluorescent microscopy.
Dual-luciferase reporter assay
To assess the effect of rRNA promoter activity by ZNF545, gastric cancer cells (AGS, MGC803) were plated on a 24-well plate and cotransfected with the human polymerase I (Pol I) luciferase reporter plasmid, pHrD-IRES-Luc, and pRL-CMV vector with pcDNA3.1-Flag-ZNF545 or pc-DNA3.1. Cells were harvested 24 h after transfection, and luciferase activities were analysed by the dual-luciferase reporter assay system (Promega). Reporter activity was normalised to the control Renilla luciferase. Experiments were performed in triplicate.
Pre-rRNA analysis by northern blot
For assay of ZNF545 overexpression, RNA was obtained from stable ZNF545-expressing MGC803 cells or control MGC803 cells. For ZNF545 knockdown assay, GES1 cells were transfected with ZNF545 small interfering RNA (siRNA) (sense, 5′-CCGUGGUUAUCAUCUUAUUdTdT-3′; antisense, 3′-dTdTGGCACCAAUAGUAGAAUAA-5′) (RiboBio, Guangzhou, China) or negative control siRNA (RiboBio) and harvested after 24 h of transfection for RNA extractions. Total RNA was isolated with TRIzol Reagent (Invitrogen). A pre-rRNA DNA probe was generated by amplifying human ribosomal DNA (rDNA) sequences from −65 to +438 using the forward primer 5′-CCCGGGGGAGGTATATCTTT-3′ and the reverse primer 5′-GACGTCACCACATCGATCAC-3′, and then gel-purified and random prime labelled with digoxigenin-11-dUTP using the DIG High Prime DNA Labelling and Detection Starter Kit II (Roche Applied Science, Indianapolis, Indiana, USA). Northern blots were performed by electrophoresis of 25 μg RNA samples in standard formaldehyde–agarose gels followed by transfer to Hybond-N+ membranes (Amersham/GE Healthcare Biosciences, Pittsburgh, Pennsylvania, USA) and UV cross-linking. Membranes were hybridised with the digoxigenin-labelled probe and detected according to the manufacturer's instructions (Roche Applied Science). Ethidium bromide staining of 28S was used as loading control.
Run-on transcription assay
For immunodetection of nascent rRNA, pulses of 5-fluorouridine (5-FU) (F5130; Sigma) were administered as described previously.5 Stable ZNF545-expressing MGC803 cells or control MGC803 cells were seeded on coverslips on a six-well plate. Pulses of 5-FU were performed by adding 5-FU to a final concentration of 2 mM in the culture medium. After 10 min, cells were fixed with 3.7% paraformaldehyde for 5 min. The incorporation of 5-FU into nascent RNA was detected with an antibody against halogenated UTP (1:50, anti-bromodeoxyuridine (BrdU) clone BU-33, B8434; Sigma) and a Texas Red-conjugated secondary antibody.
HP1β knockdown assay
MGC803 cells were cotransfected with 50 nM HP1β siRNA (sense, 5′- GGAAGGGAUUCUCAGAUGA dTdT-3′; antisense, 3′-dTdT CCUUCCCUAAGAGUCUACU-5′) (RiboBio) or control siRNA (RiboBio) and expression vector pcDNA3.1+ or pcDNA3.1-Flag-ZNF545 using lipofectamine 2000 (Invitrogen) according to the manufacturer's protocols. At 48 h after transfection, cells were harvested and cross-linked for the chromatin immunoprecipitation (ChIP) assay.
Chromatin immunoprecipitation
ChIP experiments were performed as previously described.6 Briefly, stable ZNF545-expressing MGC803 cells and control cells were cross-linked with formaldehyde for 10 min at room temperature. The cross-linking reaction was stopped by the addition of glycine, and the cells were collected for nuclear protein extraction. Chromatin from extracted cross-linked nuclei was sheared by sonication and precipitated with antibodies to Flag (F1804; Sigma-Aldrich) or trimethylated histone H3 at the Lys4 residue (H3K4me3; 07-043; Milipore, Billerica, Massachusetts, USA); immunoprecipitated protein–DNA complex was then captured with protein-G magnetic beads. Equal numbers of cells ((1–2)×107) were used for each immunoprecipitation. The same amount of non-specific IgG was used as control. After reversal of the cross-link and digestion of proteins with proteinase K, the immunoprecipitated DNA was isolated by the phenol/chloroform/isoamyl alcohol method. Levels of human rDNA were examined by real-time PCR with primer sequences from −65 to +33 (forward, 5′-CCCGGGGGAGGTATATCTTT-3′; reverse, 5′-CCAACCTCTCCGACGACA-3′). Fold enrichment was calculated relative to the background detected with non-specific IgG using the following formula: fold enrichment =2(CT of IgG - CT of antibody). The CT (cycle threshold) is defined as the number of cycles required for the fluorescent signal to cross the threshold in a real-time PCR assay. The experiment was repeated three times independently.
Coimmunoprecipitation
MGC803 cells were cotransfected with pcDNA3.1-Flag-ZNF545 and pEGFP-C1-HP1β (green fluorescent protein (GFP)-tagged mouse HP1β). After 48 h of transfection, total protein was extracted in RIPA buffer (5% sodium deoxycholate, 0.1% SDS, 10 mM Na3VO4, 50 mM NaF, 1% Nonidet P40, phosphate-buffered saline buffer, 1×protease inhibitor mixture) for 1 h on ice. Cell lysates were centrifuged and the pellet was discarded. Protein lysates were precleared by incubation with 20 μl protein G–agarose at 4°C for 1 h. After a brief centrifugation, the supernatant was transferred to a new tube. Each immunoprecipitation reaction was performed with 300 μg precleared cell lysate, 2 μg antibody to Flag and 50 μl protein G slurry for overnight incubation at 4°C. The supernatant was removed, and the protein G beads were washed in lysis buffer. Finally, immunoprecipitated protein complex was added with 2× loading buffer and boiled to denature the protein and separate it from protein G beads. The precipitated proteins were evaluated by western blot using primary antibody to GFP (Abcam, Cambridge, Massachusetts, USA).
Statistical analysis
Data are presented as mean±SD. The independent Student t test was used to compare the difference between two preselected groups. Relationships between ZNF545 methylation and clinicopathological characteristics of patients with gastric cancer were compared using Pearson's χ2 test. Univariate and multivariate Cox regression analysis was performed to assess the prognostic value of ZNF545 methylation. Overall survival in relation to methylation status was evaluated from Kaplan–Meier survival curves and the log-rank test. p<0.05 was taken as statistical significance.
Additional experimental procedures are provided in the online supplementary information.
Results
Silence or downregulation of ZNF545 by promoter methylation of ZNF545 in gastric cancer
ZNF545 was expressed in normal human gastric tissue and the normal human gastric epithelial cell line (GES1) (figure 1B). In contrast, of the 18 gastric cancer cell lines examined, mRNA expression of ZNF545 was silenced in 14 and reduced in two, leaving YCC10 and BGC823 with some expression (figure 1B). To examine the contribution of promoter methylation to the downregulation of ZNF545, its methylation status was examined by methylation-specific PCR (MSP). Promoter methylation was observed in all 16 silenced/downregulated gastric cancer cell lines, but no methylation was observed in BGC823 and slight methylation in YCC10, both of which exhibit ZNF545 expression (figure 1B). The normal gastric epithelial cell line, GES1, showed ZNF545 expression, but no promoter methylation. The ZNF545 methylation status was validated by bisulfite genome sequencing (BGS). The BGS results were consistent with those of MSP, in which dense methylation was found in methylated cell lines (AGS, MKN45 and KatoIII), but not in the unmethylated BGC823 and GES1 cell lines and normal gastric tissues (figure 1C). To test whether methylation directly mediates ZNF545 silencing, we treated four silenced cell lines (MKN28, AGS, KatoIII and SNU1) with the demethylation agent, 5-Aza-2'-deoxycytidine (5-Aza; Sigma-Aldrich). This treatment restored ZNF545 expression in all the silenced cell lines (figure 1D), indicating that the transcriptional silence of ZNF545 was mediated by promoter methylation in gastric cancer cells.

ZNF545 is inactivated by promoter methylation in gastric cancer. (A) The CpG island of ZNF545. The transcriptional start site and methylation-specific PCR (MSP) and bisulfite genomic sequencing (BGS) regions are indicated. (B) Silencing of ZNF545 in gastric cancer cell lines by promoter methylation. Methylation of ZNF545 was determined by MSP. M, methylated; U, unmethylated. Normal gastric tissue was epithelial tissue, mainly comprising epithelial cells. (C) Methylation status of the ZNF545 promoter confirmed by BGS. (D) Pharmacological demethylation with 5-Aza restored ZNF545 expression.
ZNF545 inhibits gastric cancer cell growth
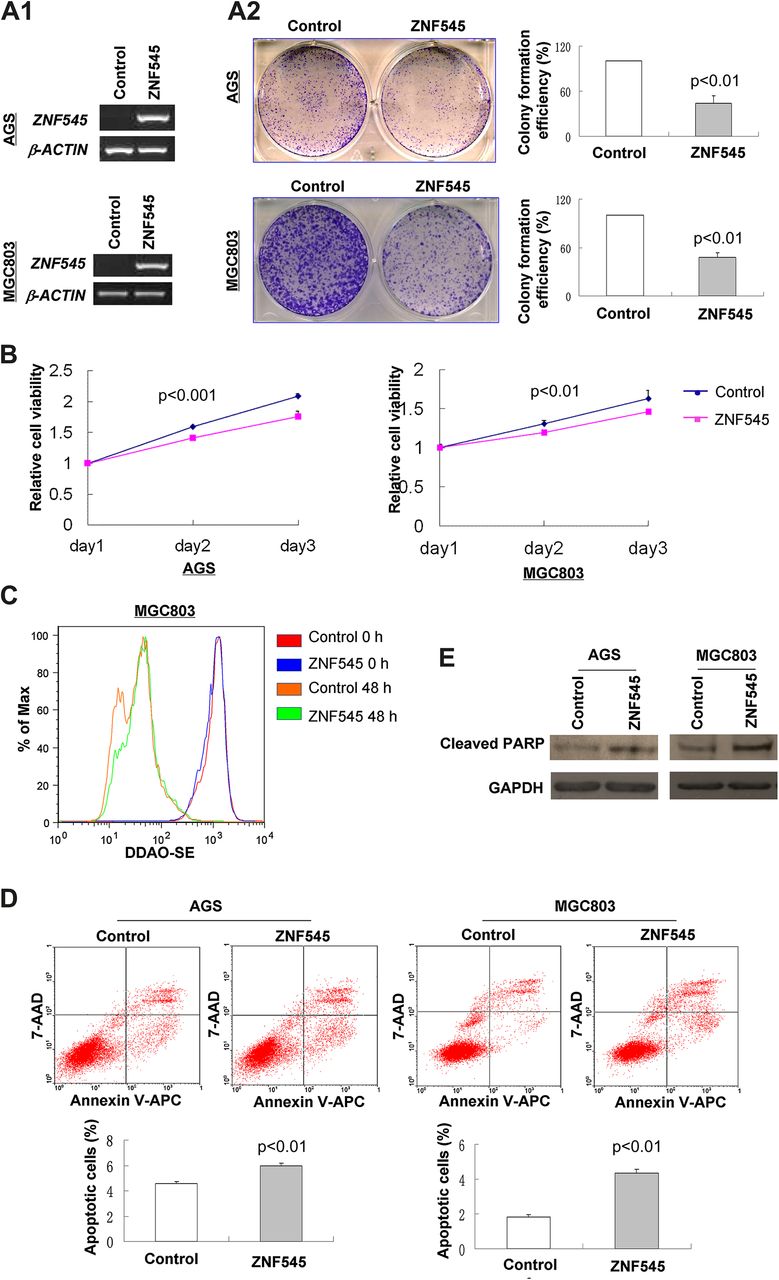
To elucidate the function of ZNF545 in gastric cancer, we examined by colony formation assay the effect of ZNF545 re-expression on growth characteristics of gastric cancer cells. ZNF545-expressing plasmid was stably transfected into AGS and MGC803 cells without ZNF545 expression (figure 2A). Re-expression of ZNF545 was confirmed by PCR analysis (figure 2A1). The colonies formed in ZNF545-transfected AGS and MGC803 cells were significantly fewer than in control vector-transfected cells (p<0.001) (figure 2A2). In addition, ectopic expression of ZNF545 caused a significant decrease in cell proliferation in AGS (p<0.001) and MGC803 (p<0.01) cells (figure 2B). The inhibitory effect on cancer cell proliferation was further confirmed by the DDAO-SE labelling assay. At 48 h after DDAO-SE labelling, the fluorescence signal of ZNF545-expressing cells was higher than that of control cells, indicating that ZNF545-expressing cells divided more slowly with less loss of fluorescence signal (figure 2C).

ZNF545 inhibited tumour cell clonogenicity and induced apoptosis of gastric cancer cells. (A) Effect of ZNF545 overexpression (A1) on colony formation in AGS and MGC803 cells (A2). (B) Effect of ZNF545 on cell viability was evaluated by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium assay in AGS and MGC803 cells. (C) Effect of ZNF545 on MGC803 cell proliferation by DDASO-SE assay. The fluorescence intensities of ZNF545-expressing cells and control cells at 0 h and 48 h were evaluated by flow cytometry after staining with DDAO-SE dye. (D) Cell apoptosis was examined by flow cytometry analysis of annexin V-APC and 7-aminoactinomycin (7-AAD) double staining. (E) Protein expression of cleaved poly(ADP-ribose) polymerase (PARP) was evaluated by western blot.
ZNF545 induces cell apoptosis
To examine the contribution of apoptosis to the observed suppression of growth by ZNF545, cell apoptosis was evaluated using double staining with annexin V-APC and 7-AAD. Ectopic expression of ZNF545 led to a significant increase in early apoptotic cells in both AGS (p<0.01) and MGC803 (p<0.01) cells compared with controls (figure 2D). In keeping with this finding, re-expression of ZNF545 significantly enhanced cleavage of poly(ADP-ribose) polymerase in stably transfected AGS and MGC803 cells compared with controls (figure 2E).
ZNF545 is localised to the nucleolus
To probe its function, the subcellular localisation of ZNF545 was examined in pcDNA3.1-Flag-ZNF545-transfected AGS and MGC803 cells using antibody to Flag. Immunofluorescent staining showed that ZNF545 was localised to the nucleolus of both cell lines (figure 3A), which is the site of ribosome biogenesis (rRNA synthesis) and is surrounded by clusters of genes that encode rRNA.7

ZNF545 inhibited rRNA transcription as a transcription repressor. (A) ZNF545 localised in the nucleolus by immunofluorescence. (B) Effect of ZNF545 on rDNA promoter activity was assessed by dual-luciferase reporter assay. (C) Levels of pre-rRNA were analysed by northern blot in response to overexpression or siRNA-mediated depletion of ZNF545 in MGC803 cells. Ethidium bromide (EtBr) staining of 28S was used as loading control. (D) De novo synthesis of pre-rRNA was monitored by in situ 5-fluorouridine (5-FU) incorporation into nascent rRNA in ZNF545-expressing and control MGC803 cells. BrdU, bromodeoxyuridine; DAPI, 4',6-diamidino-2-phenylindole.
ZNF545 inhibits rRNA transcription as a transcriptional repressor
As it is a member of the KRAB-containing zinc-finger family, ZNF545 may function as a transcriptional repressor.1 Furthermore, the above result shows that ZNF545 is localised to the nucleolus, where rRNA is transcribed.7 Thus we hypothesised that ZNF545 may play a role as a transcriptional repressor in the regulation of rRNA transcription. The dual-luciferase reporter assay was therefore performed to assess the effect of rDNA promoter activity by ZNF545. Re-expression of ZNF545 suppressed transcription on the rDNA promoter reporter plasmid in both AGS (p<0.01) and MGC803 (p<0.01) cells (figure 3B). To further confirm that ZNF545 exerts a negative influence on RNA Pol I transcription, we measured pre-rRNA synthesis in response to overexpression or siRNA-mediated depletion of ZNF545 by northern blot analysis. Pre-rRNA synthesis was greatly reduced in ZNF545-expressing MGC803 cells (figure 3C). In contrast, knockdown of ZNF545 by siRNA resulted in a clear increase in the expression of pre-rRNA compared with in control GES1 cells (figure 3C). In keeping with these findings, monitoring of de novo synthesis of pre-rRNA by 5-FU incorporation into nascent nucleolar RNA revealed that the active rRNA transcription sites, as indicated by foci of 5-FU incorporation signal, were notably decreased in ZNF545-expressing MGC803 cells compared with controls (figure 3D), suggesting that ZNF545 inhibited rRNA transcription as a transcription repressor.
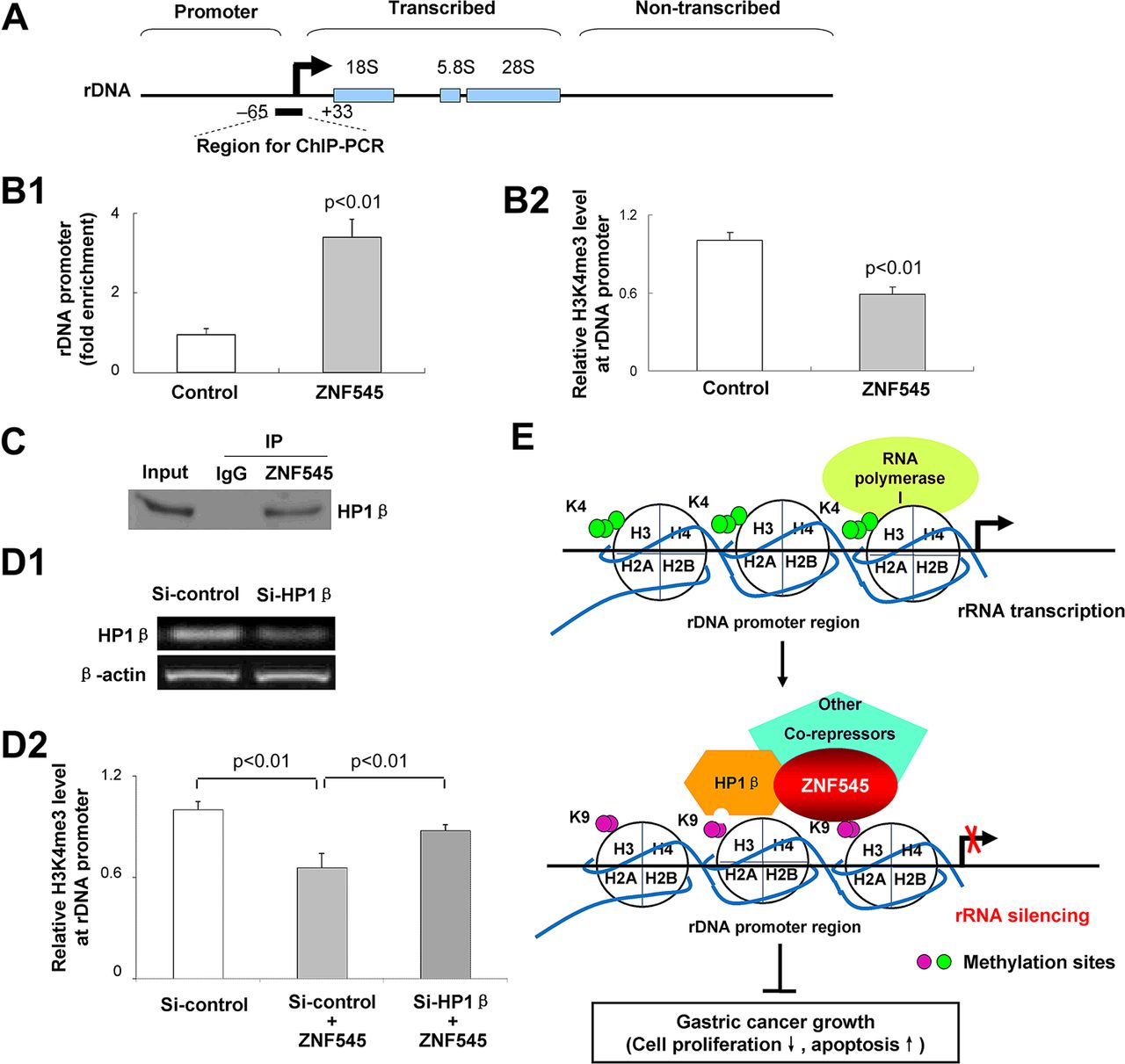
ZNF545 directly binds to rDNA promoter causing demethylation of H3K4me3
The presence of a C2H2-type zinc-finger DNA-binding domain in ZNF545 protein1 suggested that ZNF545 may bind to rDNA promoter. We thus immunoprecipitated ZNF545–DNA complex from Flag-ZNF545-expressing MGC803 cells using antibody to Flag. Quantitative ChIP-PCR was performed with specific primers targeting the rDNA promoter from −65 to +33 relative to the transcription start site as reported previously8 ,9 (figure 4A). The results of ChIP-PCR confirmed that ZNF545 bound to the core rDNA promoter region with about threefold enrichment (figure 4B1). Moreover, as regulation of rRNA transcription within the nucleolus is controlled by distinct epigenetic marks, and H3K4me3 is a critical epigenetic marker for active rRNA transcription,10 ,11 we tested the effect of ZNF545 overexpression on H3K4me3 levels in the rDNA promoter region. Our quantitative ChIP-PCR results show that ZNF545 reduced the level of H3K4me3 on the core rDNA promoter region (figure 4B2).

rRNA silencing by ZNF545 was mediated by directly binding to rDNA promoter, recruitment of the corepressor, heterochromatin protein 1β (HP1β), and loss of trimethylated histone H3 at the lysine 4 residue (H3K4me3) at the rDNA promoter locus. (A) Schematic representation of a human rDNA repeat. The primer pair for chromatin immunoprecipitation (ChIP)-PCR and their approximate positions relative to the rRNA transcription start are indicated. (B1) Fold enrichment of Flag-ZNF545 occupancy at the rDNA promoter obtained with anti-Flag versus rabbit immunoglobulin was determined by quantitative ChIP-PCR analysis using chromatin prepared from Flag-ZNF545-expressing and control MGC803 cells. (B2) The relative fold enrichment of H3K4me3 at the rDNA promoter was determined by quantitative ChIP-PCR. (C) Coimmunoprecipitation showed the physical interaction between ZNF545 and HP1β. (D) The relative fold enrichment of H3K4me3 at the rDNA promoter was examined on HP1β knockdown (D1) by quantitative ChIP-PCR (D2). (E) Proposed mechanistic model of tumour suppression by ZNF545.
ZNF545 recruits the corepressor, HP1β, for rRNA transcription repression
As the KRAB domain of KRAB-containing zinc-finger proteins serves as a scaffold for recruitment of corepressors such as HP1 to suppress RNA transcription,1 we examined by coimmunoprecipitation the protein interaction of ZNF545 and HP1. The protein complex was coimmunoprecipitated by antibody to Flag from extracts prepared from MGC803 cells cotransfected with pcDNA3.1-Flag-ZNF545 and pEGFP-C1-HP1β. HP1β was detected by western blots using antibody to GFP, indicating an association between ZNF545 and HP1β (figure 4C). Upon HP1β knockdown (figure 4D1), the H3K4me3 level on the rDNA promoter region was significantly restored in ZNF545-expressing cells compared with control siRNA-transfected ZNF545-expressing cells (figure 4D2), indicating that HP1β blockade partially relieved ZNF545-mediated rRNA repression.
Promoter methylation of ZNF545 is associated with poor survival of patients with gastric cancer
The clinical application of ZNF545 methylation was evaluated in 79 paired primary gastric cancers and in 20 healthy gastric tissue samples. Partial and dense promoter methylation of ZNF545 was detected in 51.9% (41/79) of cancer tissues and 27.0% (20/74) of adjacent non-tumour tissues (p=0.001), but none of 20 normal gastric biopsy samples (figure 5A,B). There was no correlation between the methylation status of ZNF545 and clinicopathological features of patients with gastric cancer (online supplementary tables 2 and 3). Notably, partial methylation at ZNF545 promoter was observed in most of the primary tumour samples. This is mainly because gastric tumour tissue also contains normal epithelial cells, stromal cells or infiltrating immune cells with an unmethylated part.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
ZNF545 is commonly methylated in primary gastric cancer. (A) Methylation-specific PCR (MSP) of ZNF545 promoter region in paired gastric cancers. M, methylated; U, unmethylated; N, normal; T, tumour. (B) Representative bisulfite genomic sequencing (BGS) result of ZNF545 methylation status in gastric cancer, adjacent non-tumour and normal gastric tissue samples. (C1) Kaplan–Meier survival curves show that patients with ZNF545 methylation in adjacent non-tumour tissues had poorer survival than others. (C2) Kaplan–Meier curves of patients with gastric cancer, stratified by ZNF545 methylation status, of early stage (tumour, node, metastases (TNM) I/II) and late stage (TNM III/IV).
In univariate Cox regression analysis (table 1), ZNF545 methylation in adjacent non-tumour tissues (their corresponding tumour tissues also methylated) was associated with a significantly increased risk of cancer-related death (RR 2.28; 95% CI 1.26 to 4.13; p<0.007). After adjustment for age, gender and tumour, node, metastases (TNM) stage, ZNF545 methylation in adjacent non-tumour tissues was found to predict poorer survival (RR 2.01; 95% CI 1.04 to 4.12; p=0.03) (table 2). Kaplan–Meier survival curves showed that the overall survival of patients with gastric cancer who had ZNF545 methylation in adjacent non-tumour tissues was significantly shorter than that of other patients (p=0.005, log-rank test) (figure 5C1). After further stratification by TNM stage, patients with gastric cancer and ZNF545 methylation in adjacent non-tumour tissues had significantly shorter survival in stage I/II (p<0.001) (figure 5C2), but not in stage III/IV (figure 5C2).
Univariate Cox regression analysis of potential prognostic factors for patients with gastric cancer
Multivariate Cox regression analysis of potential prognostic factors for patients with gastric cancer
Discussion
In this study, we demonstrate that ZNF545 was silenced or reduced in 16 out of 18 gastric cancer cell lines. The downregulation was shown by MSP and BGS to be due to promoter methylation. Demethylation treatment restored ZNF545 expression in all silenced cell lines with full methylation of ZNF545, suggesting that promoter methylation was the principal mechanism for the transcriptional silencing of ZNF545 in gastric cancer. As downregulation of ZNF545 was observed in gastric cancer and not in normal gastric tissues, ZNF545 may function as a potential tumour suppressor. We therefore tested the putative tumour suppressor function of ZNF545 in gastric cancer cell lines. Ectopic expression of ZNF545 in silenced gastric cancer cell lines (AGS and MGC803) dramatically suppressed clonogenicity (figure 2A), inhibited cell proliferation (figure 2B,C) and induced apoptosis (figure 2D,E), indicating for the first time that ZNF545 functions as a tumour suppressor in gastric cancer.
As a member of the KRAB-ZFP family, ZNF545 protein was shown to be localised in the nucleolus of gastric cancer cells, supporting its function as a transcriptional regulator. Thus mapping the target molecules regulated by ZNF545 will be important for understanding the molecular basis of its tumour suppressive effect. As the nucleolus is the site of rRNA synthesis, we assayed the effect of ZNF545 on rDNA promoter activity. We showed by luciferase reporter assay that rDNA promoter activity was reduced by ZNF545 in both AGS and MGC803 cell lines (figure 3B). The role of ZNF545 in inhibiting rDNA promoter activity was further elucidated and confirmed by the observation that ZNF545 inhibited pre-rRNA synthesis, as evidenced by northern blot (figure 3C) and in situ immunodetection of BrdU incorporation into nascent rRNA (figure 3D). An enlarged nucleolus with increased rRNA transcription and ribosome biogenesis has been found to be mandatory for all tumour types, as cancer cells need robust synthesis of rRNA and ribosomes to meet the high demand for rapid cell division and proliferation by supporting protein synthesis.11–14 Thus we demonstrated for the first time that the tumour-suppressive function of ZNF545 in gastric cancer appeared to be associated with the inhibition of rRNA transcription. A recent study has indicated that the MYC oncogene binds directly to rDNA loci and activates rRNA transcription to promote tumorigenesis.15 It has also been found, by isolating the RNA Pol I coactivator, UBF (upstream binding transcription factor, RNA polymerase I), from rDNA, that the tumour suppressor retinoblastoma protein (pRb) impairs RNA Pol I transcription complex formation, leading to reduced rRNA synthesis and inhibited cell growth.12 Moreover, perturbation of ribosome biogenesis can trigger cell apoptosis.12 ,16–20 Inhibition of rRNA synthesis can halt the progression of ribosome assembly and lead to nucleolar stress/structure disruption and an excess of free ribosomal proteins.17 ,18 ,20 In turn, ribosomal proteins such as L11 efflux into the nucleoplasm and bind to murine double minute 2 (MDM2), blocking MDM2-mediated p53 ubiquitination and degradation and inducing p53-mediated apoptosis.17 ,18 ,20 It is therefore likely that the inhibitory properties of ZNF545 against gastric cancer cell growth observed in this study can be explained, at least in part, by the induction of apoptosis-resultant suppression of rRNA transcription.
Having observed substantial suppression of gastric cancer cell growth by ZNF545 by inhibiting rRNA transcription, we conducted experiments designed to examine the molecular mechanisms by which ZNF545 exerts its suppressive effect on rRNA transcription. We found by quantitative ChIP-PCR that ZNF545 binds directly to the rDNA core promoter region in MGC803 cells. Moreover, ZNF545 reduced the level of H3K4me3, a transcriptionally active histone modification marker associated with relaxed chromatin structure and active transcription at the rDNA promoter.10 ,11 ,21 These results indicate that ZNF545 promoted conversion of the transcriptionally active state of chromatin at the rDNA promoter into the repressive state, and thus ZNF545 may act as a transcriptional suppressor. We further demonstrated that ZNF545 recruited the corepressor, HP1β, as demonstrated by the coimmunoprecipitation assay. In addition, HP1β knockdown partially relieved ZNF545-mediated inhibition of rRNA transcription, indicating that HP1β was an essential corepressor binding target for ZNF545 in regulating rRNA repression. It has been reported that HP1 binds to the repressive chromatin marker, methylated H3K9, which contributes to chromatin condensation and maintenance of the close structure of heterochromatin.22−24 In keeping with our findings, others have reported that KRAB-ZFP binds specifically to a gene promoter through its multiple zinc fingers, whereas its KRAB domain recruits corepressors, including HP1, to form heterochromatin and silences gene expression.1 ,24 Collectively, these findings raise the possibility that ZNF545 binds directly to rDNA promoter and recruits the corepressor HP1β, leading to decreased chromatin in an active condition at the rDNA promoter and ultimately rRNA silencing, thereby contributing to the tumour-suppressive effect (figure 4E).
To ascertain the clinical application of ZNF545 in gastric tumorigenesis in vivo, we examined by BGS promoter methylation of ZNF545 in 79 paired primary gastric cancers and 20 normal controls. ZNF545 promoter methylation was observed in 51.9% of primary gastric tumours, but not in normal gastric tissues, suggesting that methylation-mediated ZNF545 inactivation is a common event in gastric carcinogenesis. In some cases (27.0%), ZNF545 methylation was observed in both the tumours and the adjacent non-tumour tissues, although the degree and frequency of methylation were distinctly higher in the tumours than in their corresponding non-tumour tissues. Notably, among the non-tumour tissues, ZNF545 methylation was commonly detected in precancerous lesions and chronic gastritis (online supplementary table 3), suggesting that aberrant ZNF545 promoter hypermethylation occurred early in premalignant lesions of gastric mucosa and accumulated during the process of gastric carcinogenesis. Loss of ZNF545 expression by promoter methylation contributed to gastric cancer progression by facilitating cell aberrant growth and evading apoptosis. We found that ZNF545 methylation detected in adjacent non-tumour tissues could significantly predict poorer survival in patients with early stage gastric cancer (table 2 and figure 5C). This is probably because adjacent normal gastric tissues containing precancerous lesions were not removed by surgery, whereas the precancerous lesions with ZNF545 methylation are more predisposed to malignant transformation and cancer development. Thus ZNF545 methylation may be regarded as a valuable new prognostic factor for patients with early stage gastric cancer.
In conclusion, we have identified a novel functional tumour suppressor gene, ZNF545, which is inactivated by promoter methylation in gastric cancer. ZNF545 suppresses gastric cancer growth by silencing rRNA transcription through direct binding to rDNA promoter, inducing histone modification changes and recruitment of the corepressor, HP1β. ZNF545 methylation detected in adjacent non-tumour tissues was associated with shorter survival in early stages of gastric cancers. Thus ZNF545 methylation may serve as a useful biomarker for early stage gastric cancer.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
Footnotes
-
Funding This project was supported by National Basic Research Program of China (973 Program, 2010CB529305); Research Grants Council RGC CERG CUHK (473008); Group Research Scheme CUHK (3110043); CUHK Focused Investment Gant (1903026), and RFCID (10090942, 11100022).
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Clinical Research Ethics Committee of the Chinese University of Hong Kong.
-
Provenance and peer review Not commissioned; externally peer reviewed.