Article Text

Abstract
Objective Western diet is a risk factor for Crohn's disease (CD). Carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) is abnormally expressed in CD patients. This allows adherent-invasive Escherichia coli (AIEC) to colonise the gut mucosa and leads to inflammation. We assessed the effects of a high fat/high sugar (HF/HS) Western diet on gut microbiota composition, barrier integrity and susceptibility to infection in transgenic CEABAC10 mice expressing human CEACAMs.
Design Colonic microbiota composition and susceptibility of CEABAC10 mice to AIEC LF82 bacteria infection were determined in mice fed a conventional or HF/HS diet. Barrier function and inflammatory response were assessed by studying intestinal permeability, tight junction protein and mucin expression and localisation, and by determining histological score and levels of cytokine release.
Results HF/HS diet led to dysbiosis in WT and transgenic CEABAC10 mice, with a particular increase in E coli population in HF/HS-fed CEABAC10 mice. These mice showed decreased mucus layer thickness, increased intestinal permeability, induction of Nod2 and Tlr5 gene transcription, and increased TNFα secretion. These modifications led to a higher ability of AIEC bacteria to colonise the gut mucosa and to induce inflammation.
Conclusions Western diet induces changes in gut microbiota composition, alters host homeostasis and promotes AIEC gut colonisation in genetically susceptible mice. These results support the multifactorial aetiology of CD and highlight the importance of diet in CD pathogenesis.
- DIETARY - GASTROINTESTINAL INFECTIONS
- IBD BASIC RESEARCH
- INTESTINAL BARRIER FUNCTION
- GUT INFLAMMATION
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
Abnormal carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) expression allows adherent-invasive Escherichia coli (AIEC) bacteria to colonise the ileal mucosa of Crohn's disease (CD) patients, which then triggers inflammation.
-
Epidemiological studies show that Western diet, in particular its composition and fat content, is a risk factor for the development of CD.
-
Dysbiosis has been reported in many CD patients. In animal models, a high fat–high sugar diet greatly modifies gut microbiota composition.
-
Increased intestinal permeability is associated with CD patients. In animal models, a high fat diet increases intestinal permeability.
-
High fat diet induces a low-grade inflammation, for which altered gut microbiota has been suggested as the trigger.
What are the new findings?
-
Western diet leads to a shift in microbiota composition comparable with what is observed in CD patients, with an increase in the mucin-degrading bacterium Ruminococcus torques, and the group Bacteroides/Prevotella. E coli populations are particularly promoted in CEACAM-expressing mice receiving a Western diet.
-
Western diet modulates innate immune gene expression, alters intestinal permeability and decreases barrier function by affecting goblet cell number, mucin expression and mucus thickness in CEACAM-expressing mice.
-
Western diet affects the host homeostasis and gut microbiota, allowing AIEC bacteria to better colonise gut mucosa, leading to a strong release of pro-inflammatory cytokine TNFα.
How might it impact on clinical practice in the foreseeable future?
-
These data support the multifactorial aetiology of CD, which involves pathogen colonisation in a genetically predisposed patient having a Westernised lifestyle. Food intake is strongly involved in gut microbiota composition and thus changes in eating habits could decrease the proportion of invasive bacteria in the gut mucosa of susceptible CD patients.
Background and aims
Inflammatory bowel diseases (IBD), in particular Crohn's disease (CD) and ulcerative colitis, are chronic disorders of unknown aetiology that affect about 150–200 cases per 100 000 in Western countries. Several genetic, immunological and environmental factors have been incriminated.1–3 Many investigations have focused on defining which factors associated with Western lifestyle could explain the increasing prevalence of new diseases such as IBD. Among those suggested are improved hygiene, the use of antibiotics and changes in dietary habits. There is increasing evidence that new chronic diseases, such as IBD, are associated with an alteration in the intestinal microbiota, a condition called dysbiosis. Interestingly, the microbial community differs with disease subtypes; the composition and abundance of bacterial groups are different between CD and ulcerative colitis patients and between colonic-CD and ileal-CD patients,4 ,5 which suggests that the microbiota plays different roles in these disease subtypes. Although the implication of bacteria in the aetiology of CD has been demonstrated,6 ,7 it is difficult to determine whether dysbiosis is a consequence, a cause or a contributing factor of the disease. Dysbiosis in CD patients is characterised by a decrease in Firmicutes,8–14 in particular Faecalibacterium prausnitzii, and an increase in Enterobacteriaceae, especially Escherichia coli.9 ,12 ,14–17 The mucolytic species Ruminococcus torques and Ruminococcus gnavus have also been reported to be more prevalent and more abundant in CD patients.11 ,12 ,18 At a subspecies level, the adherent-invasive Escherichia coli (AIEC) pathotype has been more frequently found in ileal-CD patients than in healthy controls9 ,19–22 and has been comprehensively linked to many characteristics of CD pathogenesis, which suggests it has an aetiological role rather than being a consequence of inflammation.
Western diet, enriched in total fat, animal protein, n-6 polyunsaturated fatty acids and refined sugars, is considered as a risk factor for the development of CD.23 Interestingly, CD patients often show an accumulation of ‘creeping fat’ surrounding the intestine, which is metabolically active and induces tumor necrosis factor (TNF)α and IL-6 release.24 Diet-induced animal models developed to study obesity and metabolic disorders have shown that a high fat/high sugar (HF/HS) diet dramatically changes gut microbiota composition.25 The subsequent dysbiosis has a crucial role in increased permeability, which eventually results in low-grade inflammation and metabolic disorders.26 These observations indicate that environmental factors that influence the intestinal microbiota may trigger disease.
Carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) acts as a receptor for AIEC via type 1 pili.27 CD patients, who are more susceptible to be colonised by AIEC, especially those with ileal involvement of the disease, show an abnormal expression of CEACAMs. As observed in transgenic CEABAC10 mice expressing human CEACAMs, AIEC colonisation is CEACAM6- and type 1 pili-dependent and promotes strong intestinal inflammation.28 However, there is no evidence of the putative effects of CEACAM6 overexpression on the overall intestinal microbial community. In this work, our aims were (i) to define the microbiota of CEABAC10 mice to investigate whether the expression of human CEACAMs could lead to a dysbiosis similar to that observed in CD patients; (ii) to investigate the effects of an HF/HS Western diet on the intestinal microbiota and mucosa homeostasis in CEABAC10 mice, and (iii) to determine whether the effects of Western diet on gut microbiota and host homeostasis can facilitate AIEC colonisation in CEABAC10 mice.
Methods
Mouse diet, infection and ethics statement
Seven-week-old female mice C57BL/6 were purchased from Charles River Laboratories for reproduction with heterozygote CEABAC10 FVB/N transgenic males29 maintained under specific pathogen-free conditions in the animal care facility at the Université d'Auvergne (Clermont-Ferrand, France). Litter-mates of >8th backcross were used for experimentation. The animals were separated by sex and genotype at 4–5 weeks of age. Cages were protected with filters to avoid contamination with faecal pellets. Animal protocols were approved by the Committee for Research and Ethical Issues of the CEMEAA (Comité d’éthique en matière d'expérimentation animale, Auvergne).
Purified Diet 230 HF (Safe industry, France) was used as Western diet. It has a caloric intake of 5317 kcal/kg and a nutritional composition (in kcal/kg) of 13.1% proteins, 60.6% lipids and 26.3% carbohydrates, mainly saccharose, and thus contains high levels of fat and simple sugars. Conventional diet (Conv.) was purchased from Safe. Both diets were given for 12 weeks to 10–12-week-old mice.
The mice were orally challenged with 109 LF82 strain on three consecutive days. Bacterial persistence was evaluated at 1, 2 and 3 days postinfection in faecal samples. LF82 bacteria were enumerated by plating a serial dilution on Luria-Bertani (LB) agar containing 50 µg/ml ampicillin and 20 µg/ml erythromycin and incubated at 37°C overnight. To confirm strain identification, we verified that no ampicillin/erythromycin resistant bacteria were present in the mouse microbiota. In addition, PCR experiments using AIEC LF82 specific primers confirmed that the ampicillin and erythromycin resistant bacteria corresponded to AIEC LF82.
Microbiota diversity analysis
Tissue from the proximal colon was washed with phosphate buffer saline (PBS) and used for microbiota analysis. Genomic DNA was extracted and purified with NucleoSpin Tissue (Macherey-Nagel GmbH & Co.) in accordance with the manufacturer's instructions. To optimise bacterial DNA extraction, an overnight incubation with lysis buffer (in this condition, the tissue is completely solubilised) was performed, and the lysis temperature was increased to 95°C for Gram-positive bacteria, which are difficult to lyse. Genomic DNA was used for further molecular analysis including Eubacteria profiling by denaturing gradient gel electrophoresis, quantification of relevant bacterial groups by quantitative PCR and Eubacteria sequencing using the 454 GS FLX platform. We targeted the 16S rDNA for all these techniques to study microbial diversity in the intestinal mucosa. Primers, probes and procedures are specified in online supplementary information and online supplementary table S1.
Intestinal permeability measurement
In vivo intestinal permeability was measured at 6 weeks of diet by oral administration of fluorescein isothiocyanate FITC-dextran 4 kDa (Sigma), and further detection of fluorescence in plasma as described elsewhere.30 After sacrifice, pore-forming claudin-2 protein expression was observed histologically in colonic tissue by immunofluorescence staining as previously described.30
Quantitative PCR of host genes
PBS-washed colonic tissue contiguous to the part used for microbiota analysis was used for host gene quantification. Total RNA was isolated using TRIzol according to the manufacturer's instructions (Invitrogen). One microgram of total RNA served as a template for cDNA synthesis using RevertAid Reverse transcriptase (Invitrogen). Host genes quantified were Nod2 and toll like receptor (Tlr)5 to test innate immunity activation, Muc2 and Tff3 to study barrier function and Tbp as a reference gene for data normalisation. Specific primer pairs were designed on two different exons to span a large intronic region (see online supplementary table S2). Quantitative PCRs of cDNAs were carried out using FastStart SYBR Green (Roche) and the following programme: 95°C 5 min; 40 cycles of 95°C 20 s, 56.4°C 30 s and 72°C 20 s; 72°C 7 min; and a melting curve from 60°C to 95°C for 20 min. Samples were quantified in triplicate and normalised to the housekeeping Tbp gene, which was quantified in the same PCR plate for each gene quantification.
IgA quantification
Faecal pellets were collected at week 7 of diet and stored at −20°C until analysis. Frozen pellets were thawed out, dried for 70 min at 35°C, and homogenised with T10 basic Ultra-Turrax in 1 ml of PBS containing antiprotease cocktail (Roche) and 0.001% of sodium azide. Homogenates were centrifuged for 20 min at 1500 g (4°C) and supernatants were re-centrifuged at 1000 g for 10 min and kept at −80°C until analysis. Immunoglobulin (Ig)A were quantified in diluted supernatants (1:100) by ELISA using anti-mouse IgA Ms clone C10-3, antimouse IgA Ms-Biotine clone C10-1 and purified IgA clone M18-254 (BD Pharmingen).
Mucus immunostaining and goblet cell staining
Segments of the distal colon were fixed without washing in water-free Methanol-Carnoy's fixative. The tissue was washed twice in absolute methanol before embedding in paraffin31 and cut into 4 µm sections. The sections were de-waxed using Xylene substitute (Sigma) and hydrated. For mucus immunostaining, antigens were retrieved by microwave heating in 0.01 M citrate buffer pH6 and the background was blocked by 5% bovine serum albumin. The sections were stained with rabbit anti-MUC2 H-300 (Santa Cruz Biotechnology Inc.) diluted 1:150 in bovine serum albumin 5%. After PBS washes, sections were incubated for 1 h with a goat anti-rabbit Alexa Fluor 488-conjugated secondary antibody (Invitrogen) supplemented with Hoechst (Sigma). Slides were mounted using Mountex mounting medium (CellPath). Tissues were visualised using a confocal microscope Zeiss LSM 510 Meta (Carl Zeiss). Goblet cells were stained with Alcian blue, Periodic Acid, Schiff reactif and Hemalun, as described previously.32 Following colouration, a final dehydratation step was performed before mounting in Eukitt and observation.
Histological inflammation score and cytokine quantification
The distal part of the colon was excised and fixed in 4% paraformaldehyde, paraffin embedded, cut into 5 μm slices and stained with H&E. The histological severity of colitis was graded in a blinded fashion by a pathologist following criteria described elsewhere.30 TNFα, IL-1β and IL-12p70 released in the culture supernatant of colon specimens after 24 h of incubation at 37°C under 5% of CO2 were quantified by ELISA (R&D systems) according to the manufacturer's instructions.
Statistics
For quantitative data, unpaired Mann–Whitney U two-tailed test was used for statistical comparisons. For qualitative data, such as contingency tables, χ2 method was used. Groups designed in UniFrac analysis from pyrosequencing data were compared in relation to their relative position in the principal component analysis (PCA) plot axes (P1 and P3) with Mann–Whitney U two-tailed test. Statistical analyses were performed with GraphPad Prism V.5.01.
Results
Intestinal microbiota associated with CEABAC10 mice
The abundance and composition of colonic microbiota were compared in CEABAC10 (N=10) and wild-type (WT) (N=10) 10-week-old C57BL/6 mice. No differences were observed between WT and transgenic mice in the abundance of total bacteria (Eubacteria) associated with colonic mucosa (figure 1A) or that of Clostridium leptum, E coli and Bacteroides-Prevotella spp. Cluster analysis of Eubacteria fingerprints obtained by denaturing gradient gel electrophoresis showed that transgenic and WT mice were mixed in the dendogram (figure 1B). This indicates that CEABAC10 mice, compared with WT mice, did not have a particular composition of bacteria associated with the colonic mucosa. Firmicutes, Proteobacteria and Bacteroidetes, as determined by pyrosequencing, predominated among all samples. Only 8.4% of sequences remained unclassified. Whether these unclassified bacteria represent novel lineages remains to be determined. Some differences in abundance of bacterial taxa between WT and transgenic mice were observed but were not statistically supported. However, Firmicutes were more abundant in WT mice (45.9% vs 31.5%) and Proteobacteria more abundant in CEABAC10 mice (35.6% vs 24.8%; figure 1C). At the level of species composition, transgenic CEABAC10 mice, compared with WT mice, did not show dysbiosis (figure 1D). Altogether, these results suggest that human CEACAM expression in this mouse model is not sufficient to cause significant changes in the intestinal microbiota.

Intestinal microbiota in CEABAC10 and WT mice. The groups were composed of 10 WT and 10 CEABAC10 mature male mice, separated by genotype into two cages, all on a conventional diet. (A) Abundance of bacterial groups in colonic tissue: Quantitative PCR of 16S from total bacteria (Eubacteria), standardised with 18S from mice, used to normalise biopsy size, and other bacterial groups, standardised with respect to total bacteria abundance. Significance was measured by Mann–Whitney U two-tailed test (median and range represented). (B) Colonic bacterial composition and inter-cage variation: hierarchical distance clustering, using a curve based-Person correlation and unweighted pair-group method with arithmetic averages dendogram construction, of total bacteria 16S rDNA amplicons separated by denaturing gradient gel electrophoresis. Each type of circle indicates a different cage. (C and D) Colonic bacterial composition by 454-pyrosequencing. (C) Mean relative abundance of phyla in WT (external ring) and CEABAC10 (internal ring). (D) Principal component analysis of unweighted UniFrac distances of colonic bacteria from four WT (blue squares) and four CEABAC10 (red circles) mice.
Influence of Western diet on intestinal microbiota
HF/HS diet, given over a period of 12 weeks, led to decreased abundance of bacteria associated with the colonic mucosa in both CEABAC10 and WT mice (p<0.05; figure 2A). Comparative analysis of phyla abundance revealed imbalances in WT and transgenic mice receiving HF/HS diet, but not to a level of statistical significance (figure 2B). For example, the abundance of sequences belonging to phylum Bacteroidetes was increased in HF/HS diet-treated mice of both mice strains (22.7% of sequences in CEABAC10 HF/HS vs 16.1% in CEABAC10 Conv.; 21.5% in WT HF/HS vs 11.6% in WT Conv.; figure 2B). Proteobacteria was associated with both CEABAC10 genetic background and HF/HS diet (CEABAC10 HF/HS: 30.0%; CEABAC10 Conv.: 35.6%; WT HF/HS: 33.1%; WT Conv.: 24.8%; p=NS). In contrast, UniFrac analysis from pyrosequencing data evidenced that HF/HS diet altered species composition in both genotypes (p≤0.01), and the subsequent dysbiosis was different in WT and CEABAC10 mice (p≤0.01; figure 2C).

Influence of Western diet on intestinal microbiota composition in CEABAC10 and WT mice. The groups were composed of 13 CEABAC10 and nine WT 24-week-old male mice fed a high fat/high sugar (HF/HS) Western diet for 12 weeks (six and five mice, respectively) or on a conventional diet (Conv.) (seven and four mice, respectively). (A) Total bacterial abundance in colonic tissue: quantitative PCR of 16S from total bacteria (Eubacteria) and 18S from mice to normalise sample size. (B and C) Colonic bacterial composition by 454-pyrosequencing. (B) Relative abundance of main phyla mean values for each group are given. (C) Principal component analysis of unweighted UniFrac distances of colonic bacteria from four WT—Conv. (red circles), four CEABAC10—Conv. (orange triangles), five WT—HF/HS (green squares) and five CEABAC10—HF/HS (blue triangles) males. (D) Abundance of relevant bacterial taxa: quantitative PCR of 16S rDNA from relevant bacterial groups standardised with total bacteria. Significance was measured by Mann–Whitney U two-tailed test (median and range represented).
Quantification of some relevant bacterial groups by qPCR (figure 2D) showed that HF/HS promoted the overgrowth of Bacteroides-Prevotella spp. in both genotypes, and especially in CEABAC10 mice (p=0.035). C leptum was not significantly affected by HF/HS diet in our mice models. The two mucin-degrading species R gnavus and R torques were enriched in HF/HS treated CEABAC10 and WT mice, but only to a level of significance for R torques (p<0.05). Interestingly, E coli populations were especially increased in CEABAC10 mice receiving the HF/HS diet (p=0.008). Bifidobacterium spp., F prausnitzii and Clostridium coccoides were scarce among mucosa-associated bacteria of all mice models (data not shown).
These observations indicate that a Western diet induces dysbiosis in mucosa-associated microbiota. Microbiota composition, which is characterised in particular by a high increase in the amounts of E coli in mice expressing human CEACAMs, is different from that of WT mice not expressing human CEACAMs.
Influence of Western diet on gut homeostasis
Under conventional diet conditions, CEABAC10 mice, as in previous studies,29 ,30 had increased permeability with respect to WT mice (p=0.012). In turn, HF/HS diet itself led to an increase in intestinal permeability in WT mice (p=0.002). The combination of the two factors, CEABAC10 genotype and HF/HS diet, resulted in a significant increase in intestinal permeability with respect to the other groups (WT Conv: p<0.001; WT HF/HS: p=0.004; CEABAC Conv: p=0.034; figure 3A), confirming that host genetics and diet exert a cumulative effect. For this reason, we focused further experiments on defining the alterations in intestinal barrier function which occur in CEABAC10 mice receiving a Western diet.

Influence of Western diet on barrier function in CEABAC10 mice. (A) Intestinal permeability assessed by FITC-dextran quantification in serum 5 h after intragastric administration in WT and CEABAC10 mice with or without high fat/high sugar (HF/HS) diet. WT conventional diet (Conv.)=10, WT HF/HS=21, CEABAC10 Conv.=13, CEABAC10 HF/HS =18. (B) Immunofluorescent staining of pore-forming protein claudin-2 in two representative CEABAC10 mice (total mice analysed: Conv.=6 and HF/HS=6). (C) Relative mRNA level of Muc2 determined by RT-qPCR (5 CEABAC10 Conv. and 5 CEABAC10 HF/HS mice). (D) Immunofluorescent staining of Mucin 2 (red) in two representative CEABAC10 mice (total mice analysed: Conv.=3 and HF/HS=3). (E) Alcian blue goblet cells staining (total mice analysed: Conv.=7 and HF/HS=5). (F) Relative mRNA level of Klf4 (5 CEABAC10 Conv. and 5 CEABAC10 HF/HS mice). (G) Relative mRNA level of Tff3 (5 CEABAC10 Conv. and 5 CEABAC10 HF/HS mice).
Pore-forming tight junction protein claudin-2, which is strongly expressed in CD patients,30 was highly expressed in the colonic enterocytes of CEABAC10 mice treated with HF/HS in comparison with mice on a conventional diet (figure 3B). Western diet also altered the mucus layer. Indeed, the relative mRNA level of Muc2 gene was significantly (p=0.002) lower in the colonic mucosa of mice on an HF/HS diet (figure 3C) and, as observed by immunofluorescent staining of mucin-2, a major glycoprotein secreted by the colonic goblet cells, the mucus layer was thinner in mice on an HF/HS diet than in mice on a conventional diet (figure 3D). Staining of goblet cells by alcian blue showed that the number of goblet cells was lower in the colonic mucosa of mice on an HF/HS diet than in mice fed a conventional diet (figure 3E). In addition, the mRNA level of Klf4, which encodes a transcription factor specific to goblet cell lineage, was decreased in the mucosa of mice on an HF/HS diet (figure 3F). Altogether, these data might explain the lower quantity of mucus in mice fed an HF/HS diet. In addition, mRNA levels of Tff3 gene (trefoil factor 3, a peptide that interacts with mucins to form the mucus barrier and promotes mucosal healing) were lower in HF/HS-treated CEABAC10 mice than in CEABAC10 mice receiving a conventional diet (p=0.060; figure 3G). In addition, the levels of faecal IgA (p=0.001) in CEABAC10 mice receiving an HF/HS diet were higher than in CEABAC10 mice fed a conventional diet (figure 4A). These results indicate that Western diet given to mice expressing human CEACAMs induces alterations that lead to a thinner and less protective mucus layer. This could allow the microbiota to reach the intestinal epithelium and subsequently induce low-grade inflammation.

Influence of Western diet on immune function in CEABAC10 mice. (A) IgA amounts in faecal pellets of 17 CEABAC10 conventional diet (Conv.) and 16 CEABAC10 high fat/high sugar (HF/HS) mice. (B) Relative mRNA levels of Nod2 and Tlr5 determined by RT-qPCR (5 CEABAC10 Conv. and five CEABAC10 HF/HS mice). (C) TNFα, IL-1β and IL-12p70 levels in the colonic tissue of CEABAC10 mice (7 CEABAC10 Conv., 11 CEABAC10 HF/HS diet). Significance was measured by Mann–Whitney U two-tailed test (median and range represented).
CEABAC10 mice receiving Western diet had an altered intestinal barrier, which may have caused changes to the innate immune response. We studied certain markers of immune function that have been associated with CD patients.33 ,34 We quantified the mRNA levels of the intracellular receptor NOD2, which recognises muramyl dipeptide from the bacterial wall, and TLR5, which recognises flagellin. Transgenic mice treated with HF/HS, in comparison with those on a conventional diet, showed higher Nod2 and Tlr5 mRNA levels in the colonic tissue (p<0.05; figure 4B). These receptors play a role of nexus between antigens and NF-κB signalling and the mitogen-activated protein (MAP) kinase pathway, activation of which results in secretion of pro-inflammatory cytokines. We therefore assessed TNFα, IL-1β and IL-12p70 levels in colonic samples and observed that Western diet led to the release of TNFα, IL-1β and IL-12p70 in the colonic mucosa of CEABAC10 mice (figure 4C).
AIEC colonisation and induction of inflammation in the gut of CEABAC10 mice receiving an HF/HS diet
Transgenic mice on HF/HS diet have impaired barrier function, altered immune response and increased TNFα secretion and could therefore be more susceptible to AIEC infection. Hence, we tested the hypothesis that CEABAC10 mice treated with HF/HS would be more easily colonised by AIEC strain LF82 than those fed a conventional diet. Faecal counts of LF82 bacteria were significantly higher in CEABAC10 HF/HS-treated mice than in CEABAC10 mice fed a conventional diet at days 1 (p<0.001), 2 (p<0.001) and 3 (p=0.005) postinfection (figure 5A). This suggests that an HF/HS diet could enhance AIEC strain LF82 colonisation of the mouse gut.
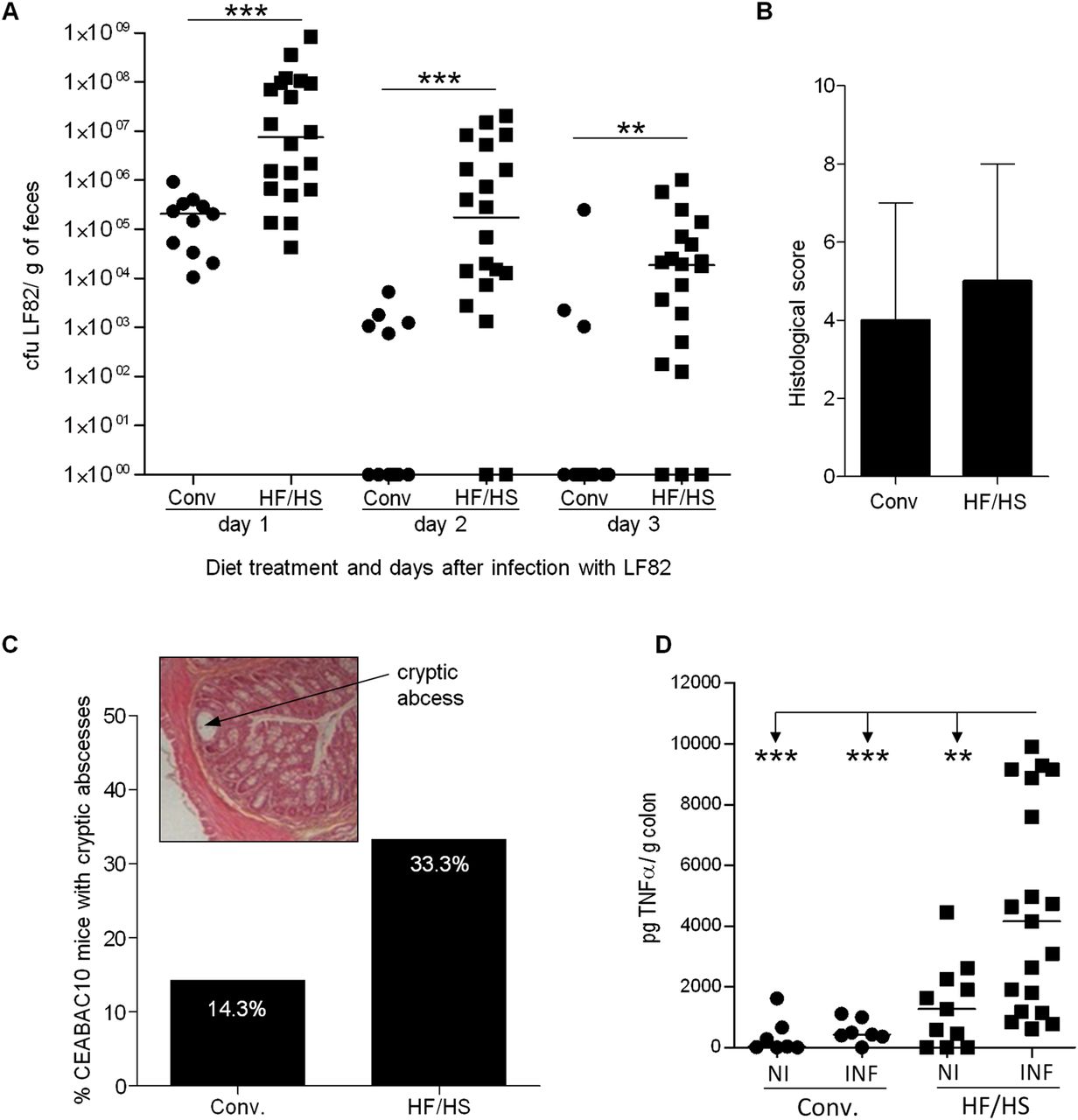
Influence of Western diet on intestinal inflammation in adherent-invasive Escherichia coli (AIEC)-CEABAC10 infected mice. (A) AIEC colonisation determined by quantification of AIEC strain LF82 in faeces after 1, 2 and 3 days postinfection by plating and colony counting in LB agar supplemented with ampicillin and erythromycin (CEABAC10 conventional diet (Conv.)=11, CEABAC10 high fat/high sugar (HF/FS)=20). (B) Histological score medians and range from colonic tissue at 3 days postinfection in CEABAC10 mice treated with conventional or HF/HS diet. (C) Frequency of mice presenting cryptic abscesses (CEABAC10 Conv.=7, CEABAC10 HF/FS=9); example of H&E/safran staining of colonic tissue sections obtained at day 3 postinfection. The arrow shows a cryptic abscess. (D) TNFα levels in the colonic tissue of LF82- infected mice (7 CEABAC10 Conv., 19 CEABAC10 and non-infected mice (7 Conv., 11 HF/HS)). Mann–Whitney U two-tailed test was applied for quantitative data.
Inflammatory signs were measured in the colonic mucosa of infected mice at day 3 postinfection. Histological examinations of CEABAC10 mice fed an HF/HS diet or conventional chow revealed similar inflammatory scores in both groups (CEABAC10 Conv.: 3.34±2.44; CEABAC10 HF/HS: 4.60±2.51; p=NS; figure 5B). Interestingly, we observed a higher percentage of infected transgenic mice presenting cryptic abscesses in animals with an HF/HS diet than in those on a conventional diet (figure 5C). These findings demonstrate that mice expressing human CEACAMs infected with AIEC LF82 develop moderate clinical symptoms of colitis and that these symptoms may be aggravated by a Western diet. Interestingly, LF82-infected HF/HS-fed CEABAC10 mice had a significant increase in TNF-α secretion compared with other groups (figure 5D), which shows that the diet and AIEC infection has a cumulative effect on colonic inflammation in CEABAC10 mice.
Discussion
We present a novel mouse model that combines diet, genetic predisposition and bacterial infection to better understand the interactions among these factors in the aetiology of CD. We used a new model of transgenic CEACAM-expressing mice (in C57BL/6 background) fed a Western-like diet, rich in fat and simple sugars, and infected with AIEC strain LF82. This model, in which inflammation is less severe than in previous ones,28 ,30 is nevertheless probably more representative of CD pathogenesis and allowed us to obtain results strongly supporting the multifactorial theory of CD aetiology. Here, we report that Western diet given to mice expressing human CEACAMs caused changes in the composition of the intestinal microbiota and led to alterations in host barrier and immunity, thereby creating a microenvironment that helped AIEC to colonise the intestinal tract and to further exacerbate the inflammatory response.
Among the factors associated with Western lifestyle, diet has a dominant role in influencing the composition of the intestinal microbiota, as previously reported in a comparative microbiota study of children from rural areas of Africa and Europe.35 In addition, diet has a clear-cut role in shaping the intestinal microbiota in animal models.25 ,26 In turn, intestinal microbial dysbiosis appears to play a major role in many clinical conditions including IBD.23 ,36 ,37 Patients with CD have increased expression of CEACAM6 on the surface of ileal mucosa.27 ,28 We show here that human CEACAM expression in CEABAC10 transgenic mice did not alter the composition of the colonic microbiota. However, Western diet highly modulated the intestinal microbial community, and the resulting dysbiosis differed between WT and CEABAC10 mice, suggesting that the genetic background may play a role in shaping gut microbiota under certain conditions. Among community-wide changes in the gut microbiota observed were lower total bacterial numbers, a greater Bacteroides-Prevotella spp. abundance and a >10-fold higher number of the mucolytic bacterium R torques associated with Western diet. We also observed that CEABAC10-treated mice on HF/HS diet had decreased Muc2 expression, which may lead to thinner mucus layers. An increase in R gnavus was also observed in the Western diet groups but not to a level of significance. To our knowledge, this alteration in mucolytic bacteria levels has not been previously observed in similar studies. Altogether, these findings indicate that the association of mucolytic bacteria with HF/HS diet warrants further investigation. The most significant result to emerge from our study is that Western diet induced dysbiosis in CEABAC10 mice, in which the E coli population was particularly increased. A higher abundance of AIEC is a trait of CD patients,21 especially those with ileal disease. AIEC are able to bind to glycosylated proteins such as CEACAMs and to colonise mucosal surfaces.27 ,38 Hence, AIEC, acting as opportunistic pathogens, benefited from the microenvironment created in Western diet-treated CEABAC10 mice. Of interest, the dysbiosis observed in Western diet-treated CEABAC10 mice was similar to that seen in CD patients,18 ,39 ,40 which lends weight to the hypothesis of a role of Western diet and genetic background in the pathogenesis of CD.
Genes involved in innate immunity were upregulated in CEABAC10 mice receiving a Western diet, as evidenced by the increased mRNA levels of Tlr5 and Nod2. These genes have been associated with CD,33 ,34 and HF diet has been reported to induce NOD2 protein expression41 and to upregulate most TLR gene expression.42 Increased expression of these pattern recognition receptors correlated with increased TNFα synthesis, indicating that antigen recognition in mice on a Western diet activates inflammatory cascade, leading to low-grade inflammation. In turn, increased IgA levels in Western diet-fed mice suggest a response of the host against bacterial populations that arise during dysbiosis, which may explain the lower total bacteria number.43 Low-grade inflammation with TNFα synthesis may have many effects on host physiology and intestinal microbiota. We demonstrate that the environment created in the intestinal mucosa of mice expressing human CEACAMs fed a Western diet facilitated the colonisation of AIEC and that AIEC benefited from the low-grade inflammation in Western diet-fed CEABAC10 mice to persist in the gut. AIEC colonisation of HF/HS-fed CEABAC10 mice correlated with increased secretion of TNFα, a pro-inflammatory cytokine strongly secreted in the intestine of CD patients.44 Western diet in interaction with microbiota is sufficient to induce intestinal inflammation mediated by TNFα synthesis via NF-κB activation.45 The amount of TNFα was greater in CEABAC10 HF/HS-infected mice than in non-infected mice, suggesting that AIEC LF82 infection was responsible for the aggravated signs of inflammation. This hypothesis is highly plausible since one mechanism of virulence of AIEC strain LF82 is its survival and replication within macrophages, which produce great amounts of TNFα.46 Cryptic abscesses were also more frequently found in infected CEABAC10 mice receiving HF/HS diet, as frequently observed in IBD patients.47
Another effect of Western diet, particularly in CEABAC10 mice, was increased intestinal permeability. CD patients exhibit increased intestinal permeability48 and the inflamed regions are characterised by an altered pattern of tight junction proteins. We observed that increased expression of the pore-forming protein Claudin-2, which is overexpressed in CD patients,30 was associated with increased intestinal permeability in CEABAC10 mice receiving an HF/HS diet. Cani et al26 demonstrated that changes in intestinal microbiota composition were the major factor involved in increased permeability in HF fed mice and triggered low-grade inflammation. In turn, this low-grade inflammation can aggravate intestinal permeability since pro-inflammatory cytokines like TNFα can drive Claudin-2 overexpression.49 Western diet also altered barrier function by decreasing Mucin-2, Klf4 and Tff3 expression as well as mucus layer thickness and goblet cell number in colonic mucosa. Low muc2 mRNA levels have been observed in patients with IBD50 and aberrant mucin synthesis has been implicated in the pathophysiology of these diseases, since reduced mucus layer thickness allows bacteria to reach the epithelium more easily. In contrast, trefoil factors are usually increased in a situation of induced inflammation to restore the damage incurred by injury,51 and thus decreased levels of TFF3 may weaken barrier function, which can lead to increased susceptibility to mucosal damage.
Our results point to a major effect of Western diet on intestinal microbiota modulation and host homeostasis disruption. We demonstrate that, in genetically susceptible hosts, Western diet makes the host more likely to be overcolonised by E coli and by AIEC when present. Successful gut colonisation by AIEC pathotype leads to the release of increased amounts of TNFα, which aggravate the inflammatory process (figure 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Model showing the effect of Western diet on microbiota composition and increased inflammation in the context of inflammatory bowel disease.
Acknowledgments
We thank Dr Abdelkrim Alloui for animal care (Animal facilities, Clermont-Ferrand, France) and Claude Darcha for histological grading (Anatomie et cytologie pathologiques, CHU, Clermont-Ferrand, France). The authors wish to acknowledge Dr Diego Morgavi (INRA, Theix) for allowing us to perform DGGE in his laboratory, Dr Yannick Bidet and his group (platform GINA, CHU Clermont-Ferrand) for performing 454-sequencing, and the CICS platform (Université Auvergne) for technical assistance with tissue preparation for microscopy analysis. We thank Jeffrey Watts for native English editing.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
MM-M and JD contributed equally to this article.
-
Contributors MMM was involved in data acquisition, statistical analysis and interpretation, and writing of the manuscript. JD was involved in data acquisition, analysis, interpretation and writing of the manuscript. ND and EB were involved in data acquisition. FR and RB performed biostatistical analysis of pyrosequencing data. ADM obtained funding and critically revised the manuscript for important intellectual content. NB obtained funding, designed and supervised the study.
-
Funding This study was supported by the Ministère de la Recherche et de la Technologie, Inserm (UMR Inserm Université d'Auvergne U1071), INRA (USC-2018) and by grants from the Association F. Aupetit (AFA), Région Auvergne (Nouveau Chercheur, 2010–2012), from ANR in the frame of ERA-NET Pathogenomics (ColiEvol project, 2009–2012).
-
Competing interests None.
-
Ethics approval This study was carried out with the approval of Committee for Research and Ethical Issues of the CEMEAA (Comité d'éthique en matière d'expérimentation animale, Auvergne).
-
Provenance and peer review Not commissioned; externally peer reviewed.