Article Text

Abstract
Objectives Long non-coding RNAs (lncRNA) have been shown to play important roles in the development and progression of cancer. However, functional lncRNAs and their downstream mechanisms are largely unknown in the molecular pathogenesis of oesophageal adenocarcinoma (EAC) and its progression.
Design lncRNAs that are abnormally upregulated in EACs were identified by RNA-sequencing analysis, followed by quantitative RT-PCR (qRTPCR) validation using tissues from 25 EAC patients. Cell biological assays in combination with small interfering RNA-mediated knockdown were performed in order to probe the functional relevance of these lncRNAs.
Results We discovered that a lncRNA, HNF1A-AS1, is markedly upregulated in human primary EACs relative to their corresponding normal oesophageal tissues (mean fold change 10.6, p<0.01). We further discovered that HNF1A-AS1 knockdown significantly inhibited cell proliferation and anchorage-independent growth, suppressed S-phase entry, and inhibited cell migration and invasion in multiple in vitro EAC models (p<0.05). A gene ontological analysis revealed that HNF1A-AS1 knockdown preferentially affected genes that are linked to assembly of chromatin and the nucleosome, a mechanism essential to cell cycle progression. The well known cancer-related lncRNA, H19, was the gene most markedly inhibited by HNF1A-AS1 knockdown. Consistent to this finding, there was a significant positive correlation between HNF1A-AS1 and H19 expression in primary EACs (p<0.01).
Conclusions We have discovered abnormal upregulation of a lncRNA, HNF1A-AS1, in human EAC. Our findings suggest that dysregulation of HNF1A-AS1 participates in oesophageal tumorigenesis, and that this participation may be mediated, at least in part, by modulation of chromatin and nucleosome assembly as well as by H19 induction.
- Oesophageal Cancer
- Cancer Genetics
- Gene Expression
Statistics from Altmetric.com
Significance of this study
-
What is already known about this subject?
-
Long non-coding RNA, or lncRNA, is an emerging class of non-protein-coding genes that are capable of regulating multiple genes involved in key cellular processes.
-
Dysregulation of some lncRNAs is functionally involved in human carcinogenesis.
-
Little is known concerning lncRNA involvement in the development and progression of oesophageal adenocarcinoma (EAC).
-
What are the new findings?
-
HNF1A-AS1, a previously unstudied lncRNA, is highly overexpressed in human primary EAC.
-
HNF1A-AS1 knockdown inhibits cell proliferation, anchorage independent growth, cell cycle progression and cell invasion/migration in multiple in vitro human EAC models.
-
HNF1A-AS1 knockdown inhibits a known cancer-related lncRNA, H19, and dysregulates chromatin-related and nucleosome assembly related genes.
-
How might it impact on clinical practice in the foreseeable future?
-
HNF1A-AS1 upregulation in oesophageal tissues may become a novel biomarker for detecting aggressive tumours with higher invasive or metastatic capacity. It may also represent a target of molecular therapies to block or reverse oesophageal tumour development.
Introduction
Oesophageal cancer is a highly lethal malignancy. Whereas the age-adjusted incidence of common cancers has decreased, the overall incidence of oesophageal adenocarcinoma (EAC) has increased rapidly over the past two decades in the USA.1 ,2 The reasons for this marked increase are not known. In addition, though alterations in oncogenes and tumour suppressor genes have been reported in EAC,3 the precise molecular mechanisms underlying EAC pathogenesis remain to be fully elucidated.
Recent research has postulated that a class of non-protein-coding RNAs (ncRNAs), known as long non-coding RNAs (lncRNAs), participates in cell fate determination and human disease pathogenesis.4 ,5 LncRNAs are non-coding RNAs greater than 200 nucleotides in length. Unlike their shorter counterparts that include microRNAs (miRNAs), mechanisms of lncRNA involvement in human disease are largely unknown. LncRNAs can control genes either locally or globally, and there are multiple means by which they can modulate downstream target genes.6 ,7 Increasing evidence suggests that lncRNAs are powerful transcriptional and post-transcriptional regulators of gene activity. Examples include lincRNA-p21 repressing TP53-induced gene transcription, PTEN ceRNAs competing with PTEN for microRNA binding, 1/2-sbsRNAs activating STAU1-mediated mRNA decay by duplexing with 3′ UTRs via Alu elements, Xist mediating the formation of transcriptionally silent nuclear compartments, and HOTAIR mediating epigenetic reprogramming via guiding Polycomb Repressive Complex occupancy to a stem cell-like state.8–,11
Accumulating data have also established the participation of lncRNAs in tumorigenesis. Examples include promotion of breast cancer cell invasion and metastasis by HOTAIR, control of breast cancer cell apoptosis by GAS5, modulation of melanoma cell apoptosis and invasion by PTEN ceRNAs and SPRY-IT1, reprogramming of induced pluripotent stem cells by RNA-RoR, and regulation of hepatocellular cancer cell growth and apoptosis by MEG3.8 ,12–,16 However, functional involvement of lncRNAs in oesophageal adenocarcinogenesis has not been extensively studied.
We recently reported that overexpression of a lncRNA contributes to the development of EAC.17 In the current study, a next-generation sequencing analysis of human oesophageal tissues uncovered marked upregulation of the lncRNA HNF1A-AS1 in EAC tissue relative to normal oesophagus. We further discovered that siRNA-mediated knockdown of HNF1A-AS1 results in diminished cell proliferation, migration, invasion and S-phase entry in EAC cells. Finally, we identified several potential downstream effectors of HNF1A-AS1. Taken together, these findings suggest that HNF1A-AS1 participates as a non-coding oncogene in oesophageal tumorigenesis.
Materials and methods
Cell culture
Primary, normal, non-immortalised oesophageal epithelial cells (HEEpiC), as well as EAC cell lines SKGT-4 and OE33, were purchased from ScienCell Research Laboratories (Carlsbad, California, USA), Sigma Chemical (St Louis, Missouri, USA), and the European Collection of Cell Culture (Porton Down, UK), respectively. The EAC cell lines FLO-1 and JH-EsoAd1 were generous gifts from David G Beer, PhD, and James R Eshleman, MD, PhD, respectively.18 ,19 SKGT-4 and OE33 were used in all cell biological assays except for invasion/migration assay. JH-EsoAd1 replaced OE33, because naïve OE33 did not invade/migrate successfully under the given experimental condition. All media were supplemented with 10% fetal bovine serum (Invitrogen, San Diego, California, USA), unless otherwise stated.
Tissues
Tissues were obtained at endoscopy performed for clinical diagnostic indications and stored in liquid nitrogen prior to total RNA extraction. All patients provided written informed consent under protocols approved by institutional review boards at the Johns Hopkins University School of Medicine, the University of Maryland School of Medicine, or the Baltimore Veterans Affairs Medical Center. All tissues were histopathologically confirmed as normal oesophagus (NE), Barrett's oesophagus (BE), or EAC. Patient descriptions are listed in online supplementary table S1.
Next-generation RNA sequencing
RNA-sequencing (RNA-seq) of oesophageal tissues was carried out using the Illumina HiSeq 2500 sequencer (single-end reads) platform as described in online supplementary methods.
Quantitative real-time polymerase chain reaction
2-step quantitative real-time polymerase chain reaction (qRT-PCR) was performed in triplicate using an oligo-dT RT primer and SYBR Green Supermix (Bio-Rad Laboratories, Hercules, California, USA) as described previously.17 Gene expression levels were normalised to β-actin expression. Online supplementary table S2 shows the primer sequences. Total RNAs from normal tissues of 20 different organs are purchased from Applied Biosystems (Foster City, California, USA; Hm total RNA survey panel kit).
Small interfering RNA transfection
Two different antihuman siRNAs per target, purchased from Life Technologies (Carlsbad, California, USA), were used: aHNF1A-AS1 siRNA1 (Cat# n265374), aHNF1A-AS1 siRNA2: (Cat# n265377), aH19 siRNA1 (Cat# n272450) and aH19 siRNA2 (Cat# n272452). Synthetic sequence-scrambled siRNA was used as a negative control siRNA. 1XPBS was used for mock-transfection. Cells were plated and cultured in growth media until cell density reached 50–60% prior to siRNA transfection using Lipofectamine RNAi MAX (Invitrogen).
Cell proliferation assays
Forty-eight hours after siRNA transfection, 2000 cells per well were seeded onto 96-well plates. After 4 h of culture (day 0), as well as at days 1, 3 and 5, cell proliferation was measured using Cell Proliferation Reagent WST-1 (Roche, Mannheim, Germany) as described previously.17
Western blotting
Cells were harvested 48 h after siRNA transfection. Lysis, electrophoresis and target protein visualisation were performed as described previously.20 We used 1:500 antihuman HNF1A goat polyclonal antibody (sc-6547; Santa Cruz Biotechnology, Santa Cruz, California, USA), and 1:10 000 antihuman β-actin mouse monoclonal antibody (A3854; Sigma–Aldrich, Bedford, Massachusetts, USA).
Soft agar colony formation and clonogenic assays
Cells were trypsinised into a single-cell suspension 48 h after transfection. For soft agar colony formation assay, 2000 cells were plated in each well of a 6-well plate which contained soft agar (Gibco, Grand Island, New York, USA) then maintained for 2–4 weeks to allow colony formation. Colonies whose diameter was greater than 150 μm were counted. Clonogenic assays were carried out as described previously.17 Colony formation ratio was calculated as ‘number of cells/initiative cell×100 (%)’.
Cell apoptosis and cell cycle analyses
Cell apoptosis was analysed 48 h after siRNA transfection using staining with Annexin V and propidium iodide (PI) as described previously.17 Cell cycle analysis was performed 48 h after siRNA transfection using PI staining according to a protocol described previously.17 Three independent experiments were performed for each assay.
Cell migration/invasion assay
Cell motility and invasiveness at 48 h after siRNA transfection were measured on Matrigel chamber plates (24-well format, BD Biosciences) as described previously.17
Microarray analysis
Microarray analysis was performed using Illumina Human-HT-12v4 BeadChips (Illumina) as described in online supplementary methods. Gene ontology (GO) term-enrichment analysis was carried out using DAVID 6.7.21
Statistical analysis
Statistical analysis was performed using SPSS software (SPSS, Chicago, Illinois, USA). Experimental results were evaluated using the two-tailed Student t test or Spearman rank correlation test. All values were expressed as mean±SD. Statistical significance was noted at p<0.05, and statistically insignificant trend was noted at p<0.1. Three independent triplicated experiments were performed for cell biological assays, unless otherwise stated.
Results
Next-generation RNA-seq analysis detects lncRNAs upregulated in EAC
In order to identify novel oncogenic lncRNAs in oesophageal adenocarcinogenesis, we carried out RNA-seq of a matched NE-BE-EAC tissue pair (full data available at GEO, Accession Number GSE48240). This RNA-seq analysis detected 6216 lncRNAs that were expressed in the EAC. Among these 6216 lncRNAs, 61 unique lncRNAs were markedly upregulated in EACs relative to NEs (fold-change >8; see online supplementary table S3). We prioritised lncRNAs that showed the highest expression in EAC (normalised copy number >20) and were sequentially upregulated during the NE-BE-EAC progression continuum (cut-off fold-change=1.5). We excluded lncRNAs with normalised copy numbers <1 in any tissues tested. This filtering process identified 3 lncRNAs (H19, HNF1A-AS1 and LOC401320). Notably, H19 is already known to be overexpressed in cancers of multiple organs, including the oesophagus.22 Therefore, the selection of H19 appears to underscore the validity of our filtering approach.
Significant HNF1A-AS1 overexpression in primary EACs and EAC cell lines is validated
We proceeded to validate differential expression of the two novel candidate lncRNAs, HNF1A-AS1 and LOC401320. qRT-PCR of these two lncRNAs in 4 EAC cell lines versus primary NE (HEEpiC) cells revealed that HNF1A-AS1 was overexpressed in all 4 EAC cell lines by 2-fold to 5-fold (figure 1A). LOC401320 was overexpressed only in OE33, JHU-EsoAd1 and SKGT4 EAC cell lines (data not shown). Therefore, we chose HNF1A-AS1 for further studies. HNF1A-AS1 is a 2455-nucleotide single-exon gene located at chromosomal band 12q24.31. Interestingly, it is also a bidirectional lncRNA corresponding to the hepatic nuclear factor 1 alpha gene (HNF1A, also known as transcription factor 1 or TCF1), whose transcriptional start site occurs approximately 5 kb 5′ to that of HNF1A-AS1 (figure 2). HNF1A-AS1 contains two Alu-like regions, as well as predicted binding sites for miRs-17, 20b, 93, 143, 644b, 4787 and 5095 (miRanda; data not shown). HNF1A-AS1 is evolutionally poorly conserved (figure 2), yet widely expressed in the aerodigestive tract (figure 1B). Subsequently, matched NE-EAC tissue pairs from 25 EAC patients (21 men and 4 women, 67.7±12.6 years of age) were interrogated for HNF1A-AS1 expression by qRT-PCR. HNF1A-AS1 expression was elevated relative to NE in the majority of EACs studied (17/25, average fold change 10.6, paired t test p value <0.01; figure 1C).
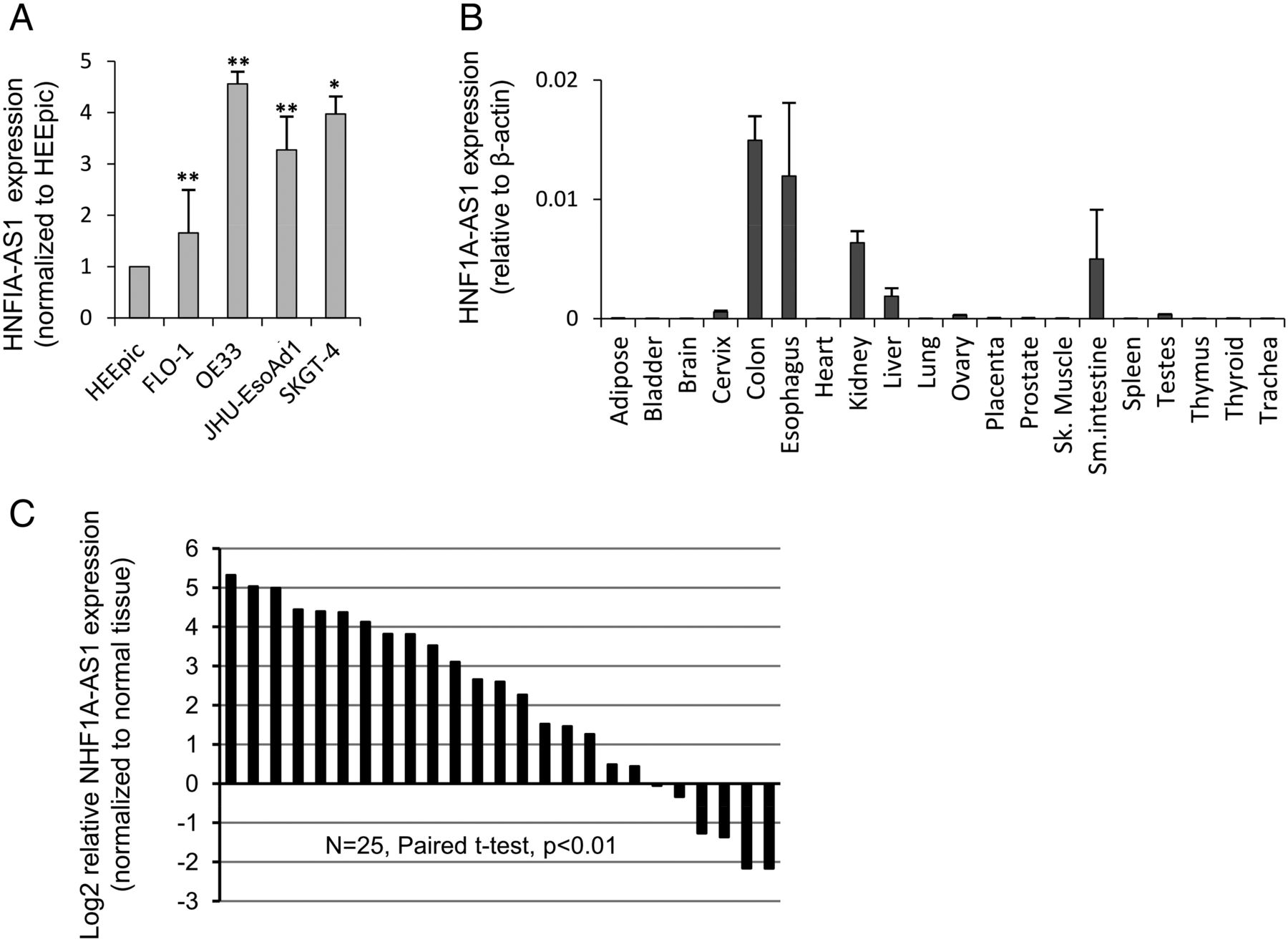
HNF1A-AS1 is highly expressed in human gastrointestinal track and is upregulated in human oesophageal adenocarcinoma (EAC). (A) Expression of HNF1A-AS1 in 4 EAC cell lines and 1 normal oesophageal epithelial cell line (HEEpiC). Expression levels are normalised to HEEpiC. (B) Expression profiling of HNF1A-AS1 across 20 normal human tissues. (C) HNF1A-AS1 expression was significantly upregulated in primary EACs relative to their corresponding normal oesophagus (NEs) in 25 EAC patients. Expression levels are shown as log2-fold change to matching NE tissue. *, p<0.05 and **, p<0.01.
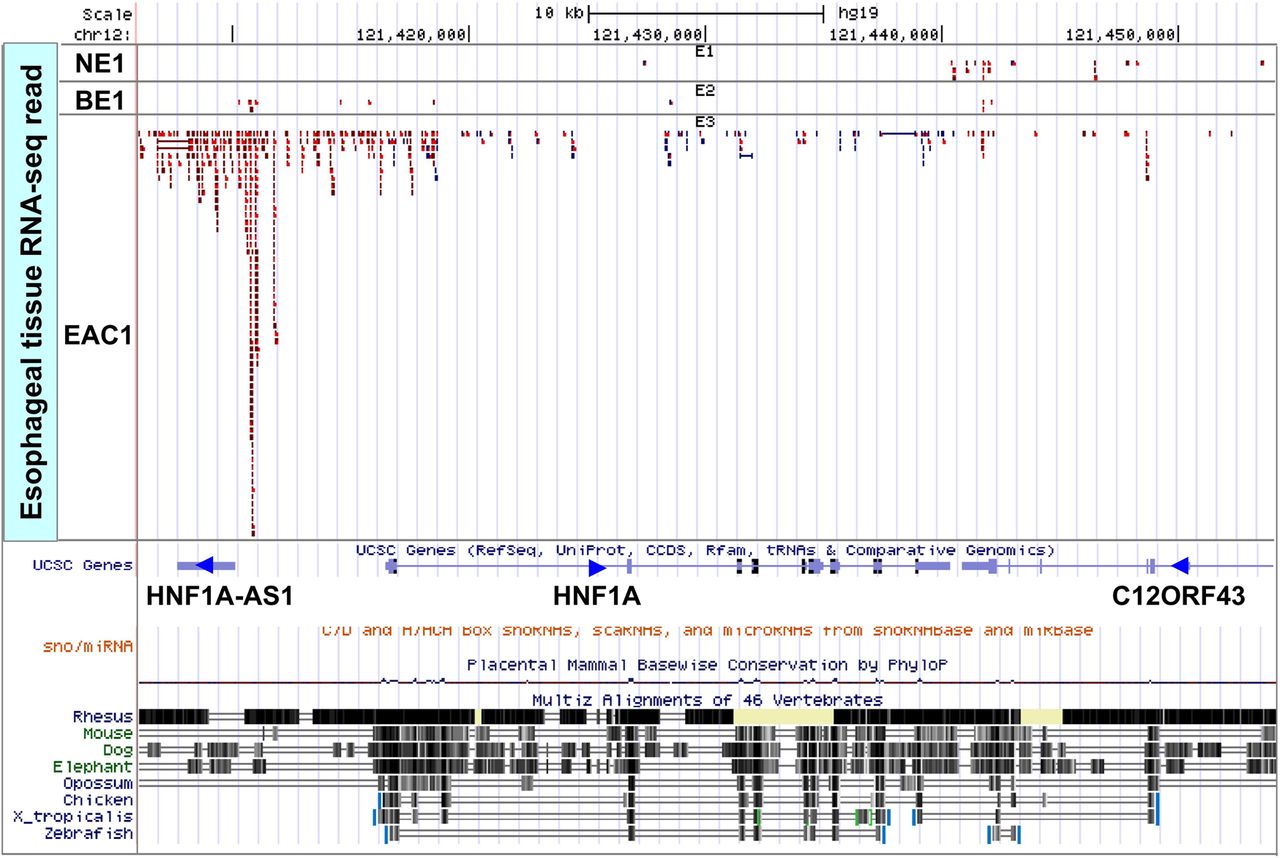
RNA-seq showed marked overexpression in oesophageal adenocarcinoma (EAC) as well as mild overexpression in Barrett's oesophagus relative to normal oesophagus for HNF1A-AS1. This illustration shows the RNA-seq data at the HNF1A-AS1 locus (12q24.3; chr12:121,406 000–121,454 000 according to the GRC37/hg19 assembly). Flanking genes did not demonstrate comparable aberrant expression, except for mild HNF1A overexpression in the analysed EAC. Blue reads corresponds to plus strand, and red reads corresponds to minus strand. Direction of gene transcription is indicated by an arrowhead.
siRNA-mediated knockdown of HNF1A-AS1 inhibits EAC cell proliferation and anchorage-independent growth
We then tested whether HNF1A-AS1 was functionally involved in EAC tumorigenesis. For this purpose, we developed an HNF1A-AS1 knockdown model using transfection of two discrete anti-HNF1A-AS1 siRNAs in EAC cell lines (see online supplementary figure S1A). WST-1 assays were used to detect the impact of HNF1A-AS1 knockdown on proliferation of the EAC cell lines, SKGT-4 and OE33. siRNA transfection-mediated HNF1A-AS1 knockdown decreased cell growth by more than 40% relative to negative control scrambled siRNA transfection at day 5 in both cell lines (p<0.05; figure 3A). Furthermore, soft agar colony formation assays revealed that anti-HNF1A-AS1 siRNA-treated cells exhibited significantly decreased anchorage-independent growth compared with the scrambled siRNA in both cell lines (by 47–69%, p<0.01; figure 3B). Colony size was also significantly smaller in HNF1A-AS1 knockdown cells than in control cells (see online supplementary figure S1B). These findings suggest that HNF1A-AS1 is involved in the promotion of EAC cell proliferation and anchorage-independent growth, two cancerous processes in epithelial cells.

Knockdown of HNF1A-AS1 inhibits proliferation and invasion of oesophageal adenocarcinoma cells. (A) WST-1 cell proliferation assays in SKGT-4 and OE33. HNF1A-AS1 knockdown induced significantly lower growth rates at day 5 (p<0.05). (B) Soft agar colony formation assays in SKGT-4 and OE33. Number of colonies was significantly reduced in HNF1A-AS1 knockdown cells than controls. (C) Cell cycle analyses in SKGT-4 and OE33. Relative to scrambled siRNA-transfected cells, HNF1A-AS1 knockdown induced significant increase in cells at G0/G1-phase. (D) Clonogenic assays in SKGT-4 and OE33. Number of colonies was significantly reduced in HNF1A-AS1 knockdown cells than controls. Representative images of colonies are shown in online supplementary figure S1C. (E) Invasion/migration assay using matrigel transwel chambers for SKGT-4. Significantly fewer cells invaded and migrated for HNF1A-AS1 knockdown cells relative to controls. *, p<0.05 and **, p<0.01.
siRNA-mediated knockdown of HNF1A-AS1 promotes G1 arrest and inhibits cell survival, but does not cause apoptosis in EAC cells
To probe potential mechanisms underlying the growth-inhibitory effects of HNF1A-AS1 knockdown, we assessed cell survival, cell cycle and apoptosis in SKGT-4 and OE33 cells. Flow cytometric cell cycle assays in these cell lines demonstrated that HNF1A-AS1 knockdown led to a significant accumulation of cells at G0/G1-phase (p<0.01) and a significant decrease in cells in S-phase (p<0.05; figure 3C). Consistent with results of cell proliferation assays, clonogenic assays revealed that HNF1A-AS1 knockdown causes a substantial reduction in cell survival in both cell lines (ie, 53–55% reduction in colony formation vs scrambled siRNA-treated cells, p<0.05; figure 3D). However, the proportion of apoptotic cells following a 48-h treatment by anti-HNF1A-AS1 versus scrambled siRNAs remained similar (see online supplementary figure S2). Thus, HNF1A-AS1-mediated promotion of EAC cell proliferation and survival appears to be mediated by modulation of the G1-S checkpoint, rather than by apoptosis.
siRNA-mediated knockdown of HNF1A-AS1 inhibits EAC cell migration/invasion
We also examined the effect of HNF1A-AS1 knockdown on EAC cell migration/invasion (figure 3E). Knockdown of HNF1A-AS1 inhibited SKGT-4 cell invasion by 30% and 70% by siRNA1 and siRNA2, respectively (p<0.05 and<0.01, respectively), and by 49% and 50% in JH-EsoAd1 (p<0.01 for both). OE33 was excluded from our analysis due to the absence of any detectable invasion/migration in naive OE33 cells. These data imply that HNF1A-AS1 involves mechanisms relevant to the promotion of EAC cell migration and/or invasion.
HNF1A, the sense-cognate gene for HNF1A-AS1, is not an HNF1A-AS1 target
The sense-cognate gene of HNF1A-AS1, hepatocyte nuclear factor 1A (HNF1A), is a key transcription factor in hepatocyte differentiation and tumorigenesis.23–,25 HNF1A is also involved in intestinal epithelial cell differentiation.23 ,26 Given the close proximity of HNF1A-AS1 to HNF1A, we hypothesised that HNF1A-AS1 could exert its biologic effects via HNF1A modulation. To better understand the relationship between HNF1A-AS1 and HNF1A, we measured HNF1A mRNA levels in the same EAC tissues examined for HNF1A-AS1. Similar to HNF1A-AS1, HNF1A was significantly upregulated in the majority of EACs (10/17, average fold change 4.7, p<0.01; figure 4A), and RNA levels of HNF1A-AS1 and HNF1A in these tissues were mildly correlated (R=0.35, p=0.08; figure 4B). HNF1A protein was also upregulated in EACs that showed HNF1A mRNA upregulation relative to their corresponding NEs (see online supplementary figure S3A). However, HNF1A-AS1 knockdown failed to significantly alter HNF1A mRNA (figure 4C) or HNF1A protein (figure 4D) expression in vitro. HNF1A knockdown also failed to significantly alter HNF1A-AS1 expression (see online supplementary figure S3B). Thus, HNF1A does not appear to be a downstream effector of HNF1A-AS1, and vice versa.

HNF1A expression is associated with HNF1A-AS1 expression in primary oesophageal adenocarcinomas (EACs), but HNF1A-AS1 knockdown doesn't induce HNF1A downregulation. (A) HNF1A mRNA expression was significantly upregulated in primary EACs relative to their corresponding normal oesophagus (NEs) in 25 EAC patients. Expression levels are shown as log2-fold change to matching NE tissue. (B) HNF1A-AS1 and HNF1A expression levels were mildly associated in 25 primary EACs (r=0.35, p=0.08). (C) HNF1A mRNA expression was not affected by HNF1A-AS1 knockdown. Expression levels are normalised to mock. (D) HNF1A protein expression was not affected by HNF1A-AS1 knockdown. *p<0.05 and **p<0.01.
HNF1A-AS1 knockdown in EAC cells dysregulates genes involved in cell cycle regulation
In a further attempt to dissect potential downstream pathways of lncRNA HNF1A-AS1, we applied microarray analysis to identify genes whose expression changed after HNF1A-AS1 knockdown in OE33 cells; 105 genes (54 downregulated and 51 upregulated) changed expression by >1.5-fold in anti-HNF1A-AS1 siRNA- versus scrambled siRNA-transfected cells (see online supplementary table S4 and figure S4A).
GO enrichment analysis revealed that chromatin/nucleosome assembly related genes were significantly over-represented among these 105 differentially expressed genes (table 1). Similarly, there was a trend toward over-representation of M-phase- and mitosis-related genes (table 1). This over-representation of M-phase-related gene downregulation may simply reflect the HNF1A-AS1 knockdown-induced decrease in M-phase cell population. However, the same conclusion is not applicable to dysregulation of chromatin/nucleosome assembly related genes. In spite of the HNF1A-AS1 knockdown-induced decrease in S-phase populations, core histone proteins, whose normal synthesis occurs during S-phase, were upregulated.27 Similarly, despite the HNF1A-AS1 knockdown-induced increase in G0/G1-phase population, two genes normally active in G1-phase were downregulated: the CENP-A chromatin assembly factor HJURP, and a p53 stabiliser HMGB.28 ,29 Taken together, these findings suggest that at least one HNF1A-AS1-responsive pathway likely involves chromatin/nucleosome assembly during the G1/S transition.
Significantly enriched gene ontology (GO) terms for genes that are differentially expressed after HNF1A-AS1 knockdown
Although some genes with known links to cancer invasion and migration were dysregulated by HNF1A-AS1 knockdown, GO enrichment analysis failed to detect over-representation of cell motility, or invasion-related pathways among these genes. This result, which conflicts with our cell biologic data, may be due in part to our use of naïve OE33 cells, which exhibit limited invasion and migration properties.
H19 expression is HNF1A-AS1-inducible in vitro and is upregulated in primary EACs
A microarray analysis revealed that downregulation of the known cancer-related lncRNA, H19, was the most substantial gene expression change in response to HNF1A-AS1 knockdown (see online supplementary figure S4A and table S4). A qRT-PCR assay verified this finding in two EAC cell lines (p<0.01; figure 5A). By contrast, H19 knockdown failed to significantly alter HNF1A-AS1 expression (figure 5B and see online supplementary figure S4B). To further validate the link between HNF1A-AS1 and H19 expression, we measured expression levels of HNF1A-AS1 and H19 in 19 matched pairs of NE and EAC tissues. As expected, H19 was upregulated in the majority of EACs (15/19, average fold change 9.3, p<0.01; figure 5C). Furthermore, HNF1A-AS1 and H19 were consistently and coordinately upregulated in EAC versus matched NE (figure 5D; R=0.78, p<0.01). Hence, coordinated upregulation of both HNF1A-AS1 and H19 occurred in EAC tissues. H19 overexpression is known to promote anchorage-independent growth and release from cell cycle arrest, while H19 suppression impedes S-phase entry and cell growth30; these phenotypes are consistent with our HNF1A-AS1 knockdown-induced phenotype. Thus, H19 may represent an important downstream effector of HNF1A-AS1, potentially mediating this lncRNA's effects on tumour growth.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
HNF1A-AS1 knockdown induces H19 lncRNA downregulation in oesophageal adenocarcinoma (EAC) cells, and H19 expression is associated with HNF1A-AS1 expression in primary EACs. (A) qRT-PCR analysis showed significant reduction of H19 in HNF1A-AS1 siRNA-treated versus scramble siRNA-treated SKGT-4 and OE33 cells. Expression levels are normalised to mock. Conversely, no significant changes were observed in HNF1A-AS1expression in H19 siRNA-treated versus scramble siRNA-treated SKGT-4 and OE33 cells (B). (C) H19 lncRNA was significantly overexpressed in EACs relative to their corresponding normal oesophagus (NEs) in EAC patients. Expression levels are shown as log2-fold change to matching NE tissue. (D) Expression levels of HNF1A-AS1 and H19 were significantly correlated in primary EACs (r=0.78, p<0.01). Expression *, p<0.05 and **, p<0.01.
In addition to H19, multiple genes known to be involved in the regulation of cell proliferation and migration were included among the 105 genes that were subjected to HNF1A-AS1 knockdown-induced dysregulation. For example, MT1E and CXCL2 were downregulated, while SERPINB5 and ZFHX3 were upregulated (see online supplementary table S4).31–,39 qRT-PCR analyses validated significant HNF1A-AS1 knockdown-induced expression changes of these genes in two different EAC cell lines (see online supplementary figure S5). Thus, these genes may also constitute potential HNF1A-AS1 downstream target pathways.
Discussion
It has now become widely accepted that mammalian genomes encode numerous lncRNAs.40 Many new functions have been ascribed to lncRNAs, and these RNA species are now attracting much interest. Nonetheless, the functional roles of most of these transcripts remain obscure. In particular, the involvement of lncRNAs in EAC pathogenesis and progression is not widely studied. The current study allowed us to make a number of key observations concerning lncRNAs in EAC. First, we identified, a previously unstudied but widely expressed (particularly in the aerodigestive tract) lncRNA, HNF1A-AS1 (GenBank accession ID: AK096009.1), as being upregulated in EAC tissues and cell lines. Second, we found that inhibition of HNF1A-AS1 expression resulted in decreased EAC cell growth, anchorage-independent growth, cell survival, S-phase entry, migration and invasion in multiple in vitro EAC models. Third, we found that nucleosome and chromatin assembly related genes, most of which are active primarily in the G1/S phase of the cell cycle, were preferentially affected by HNF1A-AS1 inhibition. Fourth, the gene most substantially downregulated by HNF1A-AS1 inhibition turned out to be another well-known cancer-related lncRNA, H19, which has been reported to exhibit effects similar to those of HNF1A-AS1. We also observed that H19 knockdown did not affect HNF1A-AS1 expression. To our knowledge, this is the first report of any lncRNA involved in the regulation of H19, or in fact, in the regulation of any other cancer-related lncRNA. Taken together, these findings suggest that HNF1A-AS1 dysregulation contributes to the development and/or progression of EAC. It is also conceivable, based on our observations, that HNF1A-AS1 downstream pathways involve H19 as well as nucleosome and chromatin assembly pathways.
BE is an established preneoplastic condition that exhibits some of EAC's molecular aberrations in less marked severity.41 In our RNA-seq analysis, HNF1A-AS1 was mildly upregulated in BE (1.9-fold, see online supplementary table S3), suggesting a link to oesophageal metaplasia. Unfortunately, we were unable to further verify this finding due to the unavailability of corresponding BEs in our cohort. However, an independent microarray dataset of 8 NE and 10 BE tissues (GEO accession: GSE34619; ref. 42) demonstrated that HNF1A-AS1 is mildly upregulated in BEs relative to NEs (2.2-fold, adjusted p<0.05), proving that mild HNF1A-AS1 upregulation marks metaplastic change in the oesophagus. HNF1A-AS1 may also serve as a potential risk stratification marker by identifying BEs that are prone to neoplastic transformation. Unfortunately, we were unable to test this hypothesis due to the unavailability of sufficient BEs matched to EACs. Additionally, HNF1A-AS1 expression may serve as a marker of tumour aggressiveness, considering its impact on in vitro cell invasion and migration. We refrained from analysing this aspect of our data, due to insufficient power of the current cohort. Further translational studies are warranted to probe the usefulness of this novel cancer-related gene, HNF1A-AS1, as a diagnostic and prognostic EAC marker.
The mechanisms controlling expression of HNF1A-AS1 remain unknown. However, HNF1A-AS1 and HNF1A, the sense-cognate gene of HNF1A-AS1, may be regulated by the same mechanisms. We make this speculation because of their observed correlated expression across most human tissues, as well as the 5′ untranscribed region shared by these two genes. Known inducers of HNF1A are HNF4A, Wnt signalling, bile acid reflux and DNA demethylation.43–,46 In support of this concept, our previous deep sequencing-based methylation analysis of a small EAC patient tissue cohort detected loss of methylation at the HNF1A-AS1 5′UTR in EACs.17 Thus, further analysis of these mechanisms, which are potentially relevant to oesophageal neoplastic progression, may provide clues to the causes of HNF1A-AS1dysregulation in oesophageal neoplasia.
We have observed that HNF1A-AS1 knockdown dysregulates chromatin/nucleosome assembly related genes in a fashion that cannot be explained by the HNF1A-AS1 knockdown-induced shift in the cell cycle. Based on these data, we propose that promotion of G1/S transition via modulation of chromatin or nucleosome assembly is a potential oncogenic mechanism for HNF1A-AS1. Indeed, the oncogenic properties of several HNF1A-AS1-inducible chromatin and nucleosome assembly related genes have been documented. For example, HJURP overexpression predicts poor prognosis in patients with breast and lung cancer, and has been suggested as an important promoter of cancer cell immortality.47 ,48 Similarly, HMGB is overexpressed in bladder cancer, and its downregulation promotes chemosensitivity of colon cancer cells.49 ,50
H19, the gene most markedly and significantly downregulated by HNF1A-AS1 knockdown, is upregulated in primary EACs.22 H19 was initially proposed as a tumour suppressor.51 However, oncogenic roles of H19, including promotion of in vivo tumorigenesis, have been documented in breast and lung cancers, where H19 overexpression has been noted.30 ,52–,54 Moreover, phenotypic changes induced by H19 inhibition and HNF1A-AS1 knockdown are similar. Therefore, H19 appears to be a potential oncogenic downstream effector of HNF1A-AS1. Our data thus provide a novel insight into potential H19 regulatory mechanisms.
The current study uncovered additional HNF1A-AS1 downstream targets with known relevance to cancer: two HNF1A-AS1-inducible genes (MT1E and CXCL2) and two HNF1A-AS1-repressible genes (SERPINB5 and ZFHX3). Metallothionein 1E (MT1E) enhances migration and invasion of glioma and bladder cancer cells.31 ,32 The chemokine ligand CXCL2 is upregulated in primary oesophageal squamous cell carcinoma and promotes cell proliferation and metastasis in vitro.33 The serine protease inhibitor, SERPINB5, commonly known as maspin, exhibits diverse tumour-suppressive functions in multiple organs.36 The transcription factor ZFHX3, also known as AT motif binding factor 1 (ATBF1), suppresses cancer cell growth and is subject to various genetic abnormalities in gastric and prostate cancers.37–,39 These genes may represent functional downstream pathways of HNF1A-AS1.
Due to the diversity and complexity of mechanisms underlying lncRNA-mediated gene regulation, it is difficult to determine direct regulatory mechanisms of lncRNAs. Binding to miRNAs or Alu-like sequences within 3′UTRs is a mechanism by which some lncRNAs modulate their target genes.8 ,9 HNF1A-AS1 contains Alu-like sequences and predicted binding sites for multiple miRs. However, genes dysregulated by HNF1A-AS1 knockdown are not the predicted targets of these miRs, except for SERPINE1 (miR-143), and do not contain HNF1A-AS1 complementary sequences, including Alu (data not shown). We have also shown that HNF1A-AS1 does not target its sense-cognate gene, HNF1A. Our observations indicate that the capacity of HNF1A-AS1 to regulate remote genes is likely to be dictated by binding to, and modulation of, protein partner(s). Further studies are needed to elucidate the precise mechanisms by which HNF1A-AS1 modulates its targets.
In summary, we have shown increased expression of the lncRNA HNF1A-AS1 in oesophageal adenocarcinogenesis. The effects of this lncRNA on cell proliferation, cell cycle regulation, migration and invasion suggest that it promotes tumorigenesis in the oesophagus. We also provide evidence that the important cancer-related lncRNA H19 may represent a downstream effector of HNF1A-AS1, thus establishing an example of mutual regulation between two cancer-related lncRNAs.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
- Data supplement 2 - Online table 1
- Data supplement 3 - Online table 2
- Data supplement 4 - Online table 3
- Data supplement 5 - Online table 4
- Data supplement 6 - Online table 5
Footnotes
-
XY and JHS are contributed equally.
-
Contributors This study was conceived by Xue Y, JHS, WJR, BC-R, MK, VKS, ES, AKV, Xiao Y, SJM and YM. All authors contributed to the study design as well as the acquisition of samples or data. The data were analysed and interpreted by Xue Y, YY, TB, AKV, SJM and YM. This article was drafted by Xue Y, TB, YY, AKV, SJM and YM, and was reviewed and approved by all authors in the current format. All who made significant contributions are listed as authors. The authors thank DGB, (University of Michigan) and JRE. (Johns Hopkins University) for their generous gifts of cell lines.
-
Funding Support for this work includes: A China Scholarship Council Award (China Scholarship Council; People's Republic of China) for XY; R01DK087454 (National Institute of Diabetes and Digestive and Kidney Diseases; USA) and R01CA146799 (National Cancer Institute; USA) for SJM; A Prevent Cancer Foundation Research Grant (Prevent Cancer Foundation; USA) for YM, SJM is an American Cancer Society Clinical Research Professor. These funding sources had no involvement in planning, execution, or reporting of this research.
-
Competing interests None.
-
Ethics approval Institutional Review Board, Johns Hopkins University.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement The RNA-seq data that are not included in this article are open to general public via data deposition to the Gene Expression Omnibus database (National Center for Biotechnology Information; USA). No personal medical information about an identifiable individual will be included in this publicly available dataset.