Article Text

Abstract
Background and aims Altered intestinal barrier is associated with immune activation and clinical symptoms in diarrhoea-predominant IBS (IBS-D). Increased mucosal antigen load may induce specific responses; however, local antibody production and its contribution to IBS aetiopathogenesis remain undefined. This study evaluated the role of humoral activity in IBS-D.
Methods A single mucosal jejunal biopsy, luminal content and blood were obtained from healthy volunteers (H; n=30) and IBS-D (n=49; Rome III criteria) participants. Intraepithelial lymphocytes, mast cells, B lymphocytes and plasma cells were studied by imaging techniques. Differential gene expression and pathway analysis were assessed by microarray and PCR techniques. Blood and luminal immunoglobulins (Igs) were quantified. Gastrointestinal symptoms, respiratory atopy and stress and depression were also recorded.
Results Patients with IBS-D showed a higher number and activation of mucosal B lymphocytes and plasma cells (p<0.05). Mast cell density was increased in patients with IBS-D (non-atopic) and in close proximity to plasma cells (p<0.05). Microarray profiling identified differential humoral activity in IBS-D, involving proliferation and activation of B lymphocytes and Igs production (p<0.001). Mucosal humoral activity was higher in IBS-D, with upregulation of germline transcripts and Ig genes (1.3-fold–1.7-fold increase; p<0.05), and increased IgG+ cells and luminal IgG compared with H (p<0.05), with no differences in blood. Biological markers of humoral activity correlated positively with bowel movements, stool form and depression.
Conclusions Enhanced small bowel humoral immunity is a distinctive feature of IBS-D. Mucosal Ig production contributes to local inflammation and clinical manifestations in IBS-D.
- B CELL
- IRRITABLE BOWEL SYNDROME
- MUCOSAL IMMUNOLOGY
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Altered intestinal epithelial barrier is associated with low-grade inflammation in patients with diarrhoea-predominant IBS (IBS-D).
Increased antibody titres against luminal bacterial components and food have been identified in sera of patients with IBS.
B cell activity has not been evaluated in the intestinal mucosa of patients with IBS.
What are the new findings?
B lymphocytes and plasma cells form part of the low-grade inflammatory infiltrate present in the jejunal mucosa of spontaneous IBS-D.
Jejunal mucosa of patients with IBS-D shows a distinctive transcriptomic profile associated with increased humoral activity and local IgG production.
Molecular markers of isotype class switch and immunoglobulin production are increased in jejunal mucosa, and not in blood, of IBS-D and are associated with major clinical manifestations.
How might it impact on clinical practice in the foreseeable future?
These findings demonstrate an active mucosal antibody response associated with clinical outcome. Therapeutic strategies directed at minimising immunoglobulin activation may be beneficial in treating IBS-D.
Introduction
Molecular and structural alterations in intestinal epithelial barrier components are associated with immunological activity and clinical symptoms in diarrhoea-predominant IBS (IBS-D).1 ,2 Abnormal barrier function facilitates the passage of potentially harmful luminal substances to the internal milieu, where the immune system plays a pivotal role in limiting antigen penetration and maintaining gut homeostasis.3 Mucosal cellular immunity is implicated in IBS pathophysiology. Although not always reported as increased in numbers,4 ,5 activated mast cells and T lymphocytes mediate epithelial disruption and leucocyte recruitment through the local release of biological mediators.2 ,6–8 By contrast, evidence of the role of humoral immunity in IBS is scant, despite the intestine being the largest immunological organ in terms of leucocyte number and antibody production.9
Effector cells of humoral activity, B cells and plasma cells play a fundamental role in both the initiation and the termination of immune responses through antigen presentation, cytokine secretion and antibody production. The main immunoglobulin (Ig) produced in the intestinal mucosa is IgA, followed by IgM, which primarily exert their function in the luminal compartment.9 The amount of IgG and IgE, with tissular effect, produced in the intestinal mucosa is much lower; however, it can rise to face luminal antigen penetration as a second line of defence.10 To increase the effectiveness of antibodies, B lymphocytes undergo class switch recombination (CSR) by changing the heavy chain of the Ig (IGH) of IgM and IgD for those of IgG, IgA or IgE, without modifying antigen specificity.11 This process requires stimulation of B cells, followed by the germline transcription of specific gene regions of the IGH and expression of the enzyme activation-induced cytidine deaminase (AID), before B cells differentiate into Ig-producing plasma cells.11 The B cell-activating factor (BAFF) contributes to B lymphocyte and plasma cell homeostasis and survival and regulates CSR and antibody production. Its effects are mediated by the surface receptors: BAFF receptor (BAFF-R), B cell maturation antigen (BCMA) and transmembrane activator-calcium modulator and cyclophilin ligand interactor (TACI).12 Although not universally, patients with IBS display increased serum antiflagellin antibodies and higher surface IgG expression on circulating B cells compared with asymptomatic controls,13 ,14 with no significant changes in B lymphocyte number within the intestinal mucosa.4 Elevated circulating IgG and IgE against food antigens15 and improvement in symptoms after exclusion diets have also been reported,16 whereas other reports did not identify a causative relationship between seric Igs and IBS.17 The role of humoral immunity in this disorder is further supported by the correlation between an allergic background and disease severity and the predominance of diarrhoea in IBS18 .
Although alterations in the function and molecular structure of the intestinal mucosal barrier have been described in IBS,2 ,19 local antibody production has not been addressed in depth in this disorder. We hypothesised that patients with IBS-D develop increased intestinal humoral activity, which contributes to mucosal inflammation and facilitates intestinal dysfunction. The aims of this study were to identify the main cellular and molecular elements responsible for Ig isotype-class switch and Ig production in both the intestine and the blood of patients with IBS-D and healthy controls and their association with clinical manifestations.
Materials and methods
Participants
Consecutive patients newly diagnosed with IBS-D using Rome III criteria20 were prospectively recruited from the gastroenterology outpatient clinic. Healthy controls (H) unmatched for sex were recruited by public advertising. All participants were ≥18 and ≤60 years of age. Prior to entering the study, candidates were asked to complete structured clinical questionnaires (to characterise digestive symptoms in patients and verify the lack of symptoms in healthy subjects), and all underwent allergy tests to rule out food allergy (see clinical assessment). A complete medical history and physical examination were carried out in all subjects. Reasonable exclusion of past episodes of infectious gastroenteritis and GI comorbidities was achieved by means of biochemical and serological analysis including antitransglutaminase antibodies and thyroid hormones in all patients. Other exclusion criteria for all subjects were pregnancy, major psychiatric and organic diseases and regular use of medication (including steroids, immunosuppressive drugs, antihistaminics and mast cell stabilisers), following the routine procedure in our laboratory.21 The study protocol was approved by the Ethics Committee of the Hospital Vall d’Hebron. Written informed consent was obtained from each participant. Inclusion and exclusion criteria for study subjects are specified in online supplementary table 1.
Clinical assessment
All participants completed daily clinical questionnaires for 10 days on (1) pain severity (by a 100-point Visual Analogue Scale),22 (2) pain frequency (number of days with pain), (3) stool frequency (maximum number of bowel movements) and (4) stool form (by the Bristol Stool Chart Score).23 Dyspepsia was also evaluated following the Rome III criteria.24 Background stress and depression levels were assessed using the validated Spanish versions of the Modified Social Readjustment Scale of Holmes–Rahe,25 the Perceived Stress Scale of Cohen26 and the Beck's Depression Inventory27. Two sets of skin prick tests (SPT) were performed with 22 common foodstuffs and 12 pneumo-allergens (Laboratorios Leti, Barcelona, Spain), using histamine and saline as positive and negative controls, respectively. Positivity was defined by skin wheal reaction (diameter >3 mm) to at least one allergen.28 Candidates with either positivity to foodstuff by SPT or clinical history consistent with food allergy (digestive and/or extra-digestive symptoms associated with exposure to certain food components) were excluded. Respiratory atopy was identified by positivity to pneumo-allergens plus a clinical history of asthma, rhinitis or eczema, and participants (both H and IBS-D) were classified as respiratory atopic (A) or non-atopic (NA).
Biological samples
Intestinal fluid collection and jejunal biopsy
After an overnight fast, a Watson's capsule (with an attached 3 mm diameter aspiration tube) was orally inserted under fluoroscopic control into the proximal jejunum, 5 cm distal to Treitz's angle. Jejunal fluid (4–5 mL) was obtained by gentle aspiration, snap frozen and stored at −80°C until analysed. A single mucosal biopsy per participant was obtained by suction with a 50 mL syringe. Biopsy was immediately split into two similar pieces with a sterile scalpel. One fragment was fixed in formalin and embedded in paraffin for further microscopic examination. The remaining fragment was placed either in RNAse-free tubes containing 500 μL of RNA Later Solution (Ambion, Madrid, Spain) and stored at −80°C until processed for RNA isolation or in fixative buffer (2.5% glutaraldehyde, 2% paraformaldehyde in 0.1 M phosphate buffer) for further transmission electron microscopy analysis.
Peripheral blood
Blood was collected directly into PAXgene Blood RNA tubes (Preanalytix/Qiagen/BD) and stored at −40°C for further RNA isolation and into plastic tubes (Plastic Serum BD Hemogard) for the obtention of serum by centrifugation after clot formation and stored at −40°C until analysed for Ig quantification.
Study design
Clinical assessment, jejunal mucosal biopsy and luminal content and peripheral blood were obtained from all participants. Mucosal leucocytes were quantified in all tissue specimens by immunohistochemistry. Due to limited biopsy size, the following analyses were performed in randomly selected subgroups of participants: (a) gene expression by microarray, (b) gene expression by PCR analysis and (c) plasma cell counts and ultrastructure by transmission electron microscopy. Immunoglobulins were quantified in the jejunal content and blood (see online supplementary table 2). Biopsy samples were randomised into three types of analysis (microarray, gene expression and electron microscopy) applying a standard method, as previously used1 ,2 (linear congruential generator algorithm; see online supplementary material). All samples were codified and blindly analysed by the study researchers (MV, AMG-C, IT).
Analytical procedures
Immunohistochemistry and immunofluorescence
Following standard procedures, tissue sections (4 mm) were incubated with antihuman CD117, CD3 or CD20 for mast cells, intraepithelial T lymphocyte (IEL) or B lymphocyte identification, respectively, following incubation with a secondary antibody (see online supplementary table 3) and developed with the Vectastain ABC kit (Vector Laboratories, UK). Positive cells in the mucosa were counted at high magnification (400×) in at least eight non-overlapping fields, and results expressed as the number of positive cells per high-power field (hpf) or per 100 epithelial cells. IgG+ cells and IgA+ cells in the lamina propria were identified after incubation with conjugated antibodies (see online supplementary table 3) following standard procedures. Positive cells were quantified and results expressed as the number of cells per mm2 using CellSens-V.1.7 software (Olympus).
Transmission electron microscopy
Tissue fragments were postfixed, dehydrated and infiltrated in Epon's resin, polymerised and contrasted. Ultrathin sections were examined using a Hitachi H-1400 microscope at 75 kV equipped with a MegaView III camera (Soft Imaging System). Plasma cells were quantified in at least 60 non-overlapping fields (2500 μm2 area) and results expressed as the number of cells per mm2. The distance between plasma cells and mast cells (membrane to membrane, in μm) was calculated with the same imaging software. The percentage of plasma cells in close proximity (<5 μm) to mast cells with respect to the total number of mast cells was also calculated.
RNA isolation, microarray analysis and cDNA synthesis
Jejunal mRNA was isolated using the RNAeasy Mini Kit and blood mRNA using the PAXgene Blood RNA Kit (Qiagen), following the manufacturer's instructions. Microarray technology (Affymetrix human gene 1.1 ST) was performed on randomly selected mucosal biopsies. Differentially expressed genes (p<0.05), with a mean fold change of ≤0.7 and ≥1.4, were submitted to pathway and network analysis using the Ingenuity Pathway Analysis methodology (IPA 7.0, Ingenuity Systems), as previously described.1 For network analysis, IPA provides a score according to the fit of supplied genes and the list of biological functions involved. A score >3 (p<0.001) indicates a >99.9% confidence that a focus gene network was not generated by chance alone (see online supplementary material). cDNA synthesis was performed using 1 μg of total RNA with the High Capacity Reverse Transcription Reagents Kit (Applied Biosystems, Madrid, Spain).
PCR and quantitative real-time PCR analysis
The expression of AID, germline transcripts (GLTs) from μGLT, γ3GLT, γ1GLT, αGLT, γ2GLT, γ4GLT εGLT and IGH for IgE (IGHE) were amplified with validated primer sets.29 ,30 Primers for IGHM, IGHG3, IGHG1, IGHGA, IGHG2 and IGHG4 were designed and validated in our laboratory (see online supplementary table 4). PCR reactions were performed as previously described.30 Integrity and cDNA load were monitored by endogenous control gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) amplification. All PCR products were gel-extracted and sequenced to confirm their identity with the corresponding sequence present in the GeneBank database (data not shown). Gene expression was visualised on 1.5% agarose gels and quantified by densitometric analysis using ImageJ software (National Institutes of Health; http://rsb.info.nih.gov.ij/). Expression of BAFF-R, BCMA and TACI as genes involved in B cell maturation and activation, CSR and plasma cell survival12 was performed by real-time PCR on an ABI PRISM 7500 FAST Sequence Detection System (Applied Biosystems, Spain) using validated TaqMan Gene Expression Assays, and the human 18S subunit ribosomal RNA gene as the endogenous control (Applied Biosystems). Each sample, including distilled water as negative control, was run in triplicate and data were analysed by the 2-ΔΔCt method.31 The expression of each gene in each subject was normalised to an endogenous gene with constant expression: GAPDH for PCR and 18S for real-time PCR. In the H group, the mean of these normalised values was then calculated, and individual data (both in H and IBS-D) were expressed as the proportion (fold change) with respect to this reference value.
Immunoglobulin quantification
Jejunal content and serum samples were assayed for Ig quantification by an ELISA technique for IgG, IgM, IgE (Bethyl Laboratories, Montgomery, Texas, USA) and IgA (Immundiagnostik, Germany) according to the manufacturer's instructions. Prior to analysis, a protease inhibitor cocktail (Sigma, Spain) was added to intestinal fluid samples to minimise protease activity.
Statistical analysis
Data are expressed as median (range) or mean±SD. Normality of data distribution was tested by the Kolmogorov–Smirnov test. Normally distributed parametric data were compared by the unpaired Student's t test (two-tailed); otherwise, the Mann–Whitney U test was used. The χ2 test was used for comparison of frequencies of GLTs between groups. Two-way analysis of variance (ANOVA) was used to test the effects of IBS-D/H and atopy, as well as for IBS-D/H and sex, on each clinical and biological response (when appropriate). ANOVAs were followed by post hoc tests. No power calculation was performed in this proof-of-concept study. p Values ≤0.05 were considered significant and were adjusted for multiple comparisons using the Benjamini and Hochberg method32 (application of corrections indicated in figure legends). Relationships between clinical and biological variables were assessed by Spearman's rs correlation. Statistical analysis was performed using GraphPad Prism 5.0 (La Jolla, California, USA) and the R statistical software.33
Results
Study population
Thirty H and 49 IBS-D subjects were included in the study. There were no differences in age or atopy frequency among participants; however, the number of women in IBS-D was higher than in H (p<0.05). Patients with IBS-D exhibited significantly greater depression and perceived stress scores than H subjects (table 1). In the IBS-D group, 63% had dyspepsia, with similar frequency and intensity of abdominal pain, number of bowel movements and stool form than non-dyspeptic patients with IBS-D (see online supplementary table 2 for individual clinical data). No differences in clinical characteristics existed in the subgroups used for different biological assessments (data not shown).
Clinical and demographic characteristics of participants
Differential mucosal transcriptome identifies humoral activity in IBS-D
Patients with IBS-D (n=13) displayed a distinctive mucosal transcriptome compared with H (n=8). Microarray analysis identified 185 differentially expressed genes, of which 171 were eligible for gene interaction and biological function analysis. IPA identified nine significant networks, mainly related to cellular development, growth and proliferation. Similar to our previous study, molecules related to immune activation, cell growth and signalling, and GI disease were identified.1 Molecules belonging to the networks, and their biological functions, are included in the online supplementary material. Interestingly, the second top network (score 32, focus molecules 18) recognised humoral response as one of the main distinctive categories, according to gene interaction (p<0.01). Genes in this network accounted specifically for maturation, proliferation, quantity, function and activation of B lymphocytes, as well as the quantity of Ig (figure 1). For microarray validation, IGH genes and genes related to B cell maturation and activation were further quantified.

Functional analysis of mucosal transcriptional signature in diarrhoea-predominant IBS (IBS-D) versus H. (A) Relationships between differentially expressed genes in the second highest scored network and related biological functions (Fx), as analysed by the Ingenuity Pathway Analysis (IPA) application. Node (gene) and edge (gene relationship) symbols are described. The intensity of the node colour indicates the degree of upregulation (red) or downregulation (green). Genes in uncoloured nodes were not identified as differentially expressed and were integrated into the computationally generated networks on the basis of the evidence stored in the IPA knowledge memory indicating relevance for this network. (B) List of categories and function annotation of genes identified in the network for its role in immunological humoral activity. p Values are indicated. p Value adjusted (false discovery rate): **<0.01 and ***<0.001.
Mucosal cell counts and activation of B lymphocytes and plasma cells in IBS-D
Villous architecture was well preserved in all specimens, no lymphoid follicles, significant lymphoplasmacytic infiltrate in lamina propria and/or IELs were detected under routine histopathological study. Specific staining revealed similar numbers of IELs (IBS-D: 19.9±8.6 vs H: 20.6±10.1 cells/100 enterocytes) between groups, with higher values in IBS-D versus H when NA subjects were considered, although statistical significance was not reached (see online supplementary figure 1). Mucosal B lymphocyte density was higher in patients with IBS-D (IBS-D: 3.0±1.8 vs H: 1.6±1.1 cells/hpf; p=0.014) and distributed along the crypts in the lamina propria (figure 2A). Mucosal plasma cells were also increased in IBS-D (IBS-D: 1413±1058 vs H: 335±278 plasma cells/mm2; p=0.026; figure 2A). Mucosal mast cells were similar in numbers in IBS-D (24.8±11.3 cells/hpf) and H (23.7±13.6 cells/hpf); considering the subgroups of NA participants, mast cell density was higher in IBS-D than in H (IBS-D: 28.4±11.4 vs H: 18.1±6.7 cells/hpf; p=0.019), as previously described.21 Moreover, the distance from plasma cells to mast cells in jejunal mucosa was significantly lower in the IBS-D group (IBS-D: 0.67±0.39 vs H: 1.72±1.05 μm; p=0.009; figure 2B), and the percentage of plasma cells located within 5 μm of mast cells was also higher in IBS-D than in H (H: 4.2±5.5 vs IBS-D: 16.5±18.8%; p=0.039). No association with atopy or sex was identified. Furthermore, quantitative analysis of BAFF-R, BCMA and TACI gene expression identified significant upregulation in the intestinal mucosa of patients with IBS-D (1.81-fold, 1.70-fold and a 1.62-fold increase, respectively; p≤0.05) compared with H (figure 2C) and not in blood (data not shown). Moreover, a positive correlation was detected in mucosal expression of BCMA and BAFF-R (r=0.778; p<0.0001), BAFF-R and TACI (r=0.773; p<0.0001) and BCMA and TACI (r=0.860; p<0.0001). No difference in cell distribution or receptor expression was associated with atopy or sex.

Density and activation of mucosal B cells, plasma cells and mast cells in H and diarrhoea-predominant IBS (IBS-D) groups. (A) Number of B lymphocytes and plasma cells in H and IBS-D groups and representative images of CD20-positive staining in the lamina propria. The insert in each image shows positive staining. (B) Number of mast cells in H and IBS-D groups (A, atopic; NA, non-atopic), distance from mast cells to plasma cells within the lamina propria in H and IBS-D and representative electron micrograph image of plasma cells and a mast cell below the epithelium. Large arrows indicate endoplasmic reticulum in plasma cells; small arrows indicate piecemeal degranulation of mast cells. Ep, epithelium; PC, plasma cell; MC, mast cell. Bar, 2 μm. (C) Quantitative gene expression of B cell-activating factor receptor (BAFF-R), B cell maturation antigen (BCMA) and transmembrane activator-calcium modulator and cyclophilin ligand interactor (TACI) in IBS-D and H groups. Dark and open symbols indicate respiratory A and NA participants, respectively. Individual values represent the fold change with respect to the average in the H group. The 18S was used as a house-keeping gene. p Values are indicated. p Value adjusted (false discovery rate): *<0.05 and **<0.01.
Increased mucosal isotype class switching activity and IGH expression in IBS-D
GLT expression is essential for CSR as it is the first molecular step in the commitment of B cells to Ig synthesis.11 In jejunal mucosa, the expression of μGLT, γ3GLT, γ1GLT and γ2GLT and the frequency of γ4GLT expression were higher in IBS-D than in H subjects. By contrast, in blood, only μGLT and γ1GLT were increased in IBS-D compared with H (table 2 and figure 3). We next analysed the transcription of IGH and identified significantly enhanced expression of IGHG1, IGHG2, IGHG3, IGHG4 and IGHE in jejunal mucosa of IBS-D compared with H. No differences were detected in blood between groups; only IGHG2 was lower in IBS-D samples (table 2, see online supplementary figure 2). AID was expressed only in mucosal samples, with no differences between groups. ANOVA analysis revealed differential expression due to atopy, with the NA IBS-D subgroup having increased μGLT, γ3GLT, εGLT, IGHG3 and IGHG1 expression compared with patients with atopic IBS-D (figure 3 and see online supplementary figure 2). Sex did not affect the expression of CSR molecular markers and Ig production.
Expression of GLT and IGH in jejunal mucosa biopsies and peripheral blood of H and IBS-D participants

Analysis of germline transcript (GLT) and activation-induced cytidine deaminase (AID) gene expression in H and diarrhoea-predominant IBS (IBS-D) groups. Representative image of GLT and AID expression in participants classified as respiratory atopic and non-atopic. Amplicons were identified by expected size and confirmed after gel purification and sequencing (data not shown). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a house-keeping gene. Lane (−) corresponds to PCR negative control (distilled water).
Increased mucosal IgG+ cells and IgG production in IBS-D
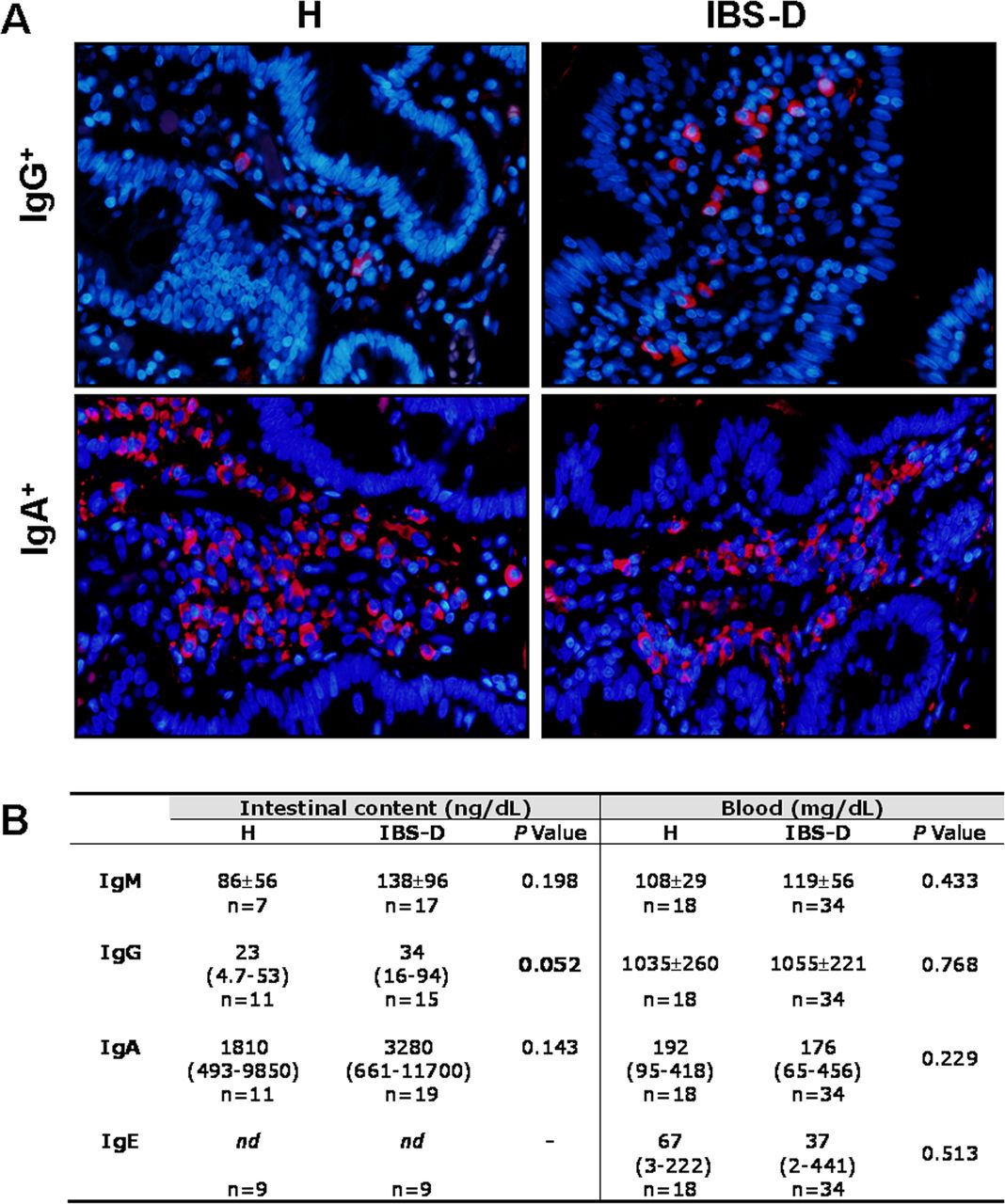
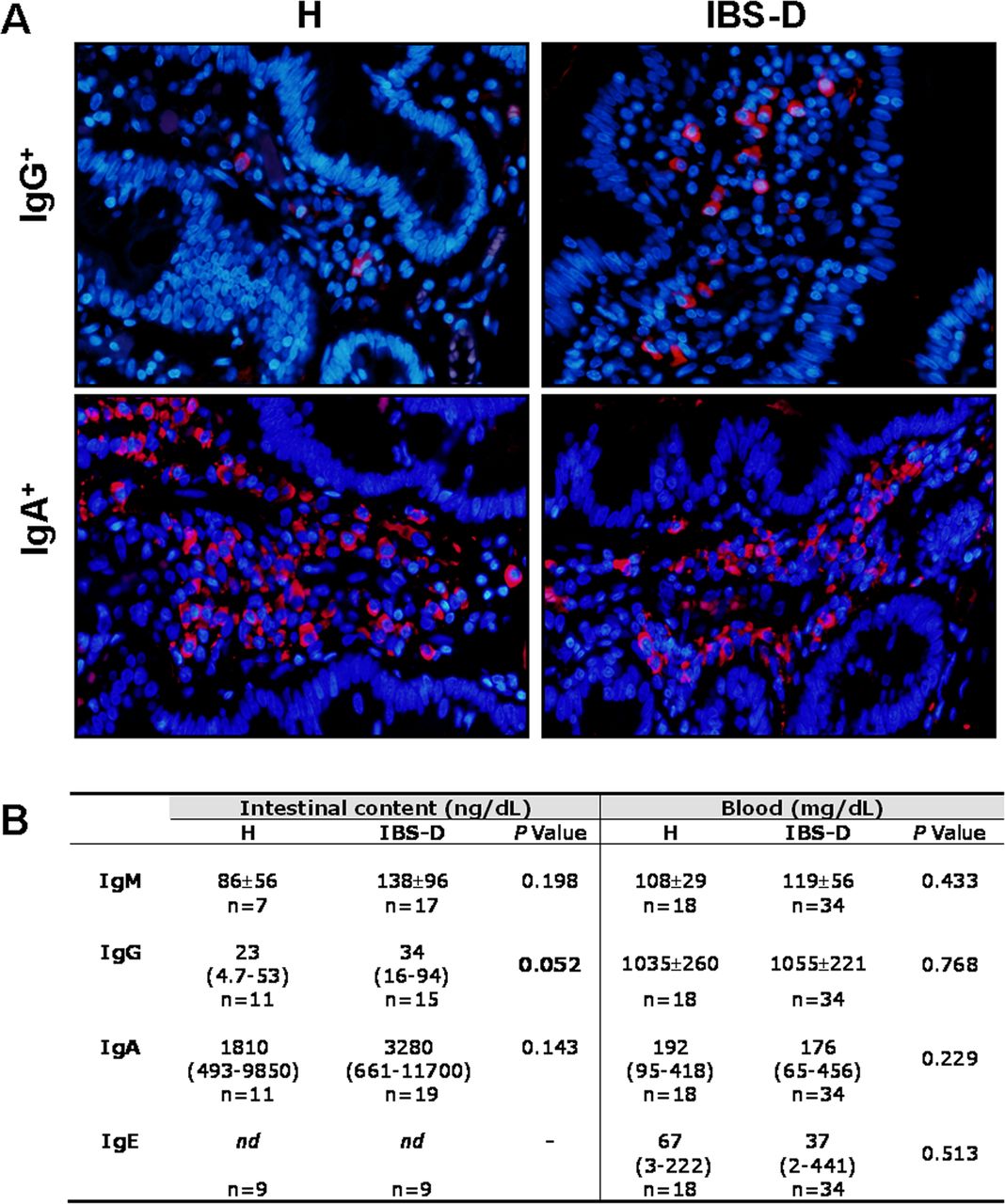
The final step of CSR is antibody production by plasma cells. The number of mucosal IgG+ cells was increased in IBS-D (613±183 cells/mm2) when compared with that in H (250±157 cells/mm2; p=0.004), whereas IgA+ cells were similar in both groups (1940±512 and 1914±363 cells/mm2, respectively). In jejunal content, IgA and IgM concentrations were higher in IBS-D than in H, although not reaching statistical significance, and IgE was below the detection limit. The luminal concentration of IgG was higher in IBS-D than in H, with no differences regarding atopy or sex. Serum Igs concentration were equivalent to reference values in all subjects, with no differences between groups (figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mucosal immunoglobulin (Ig) production in H and diarrhoea-predominant IBS (IBS-D) groups. (A) Representative images of IgG+ cells and IgA+ cells in the lamina propria of H and IBS-D subjects. Positive cells are visualised in red (Ig) and nucleus staining in blue (4’,6-diamidino-2-phenylindole). (B) Immunoglobulin concentration in the intestinal content and in blood of H and IBS-D participants. nd, not detected. p Values considered significant are shown in bold.
Clinical correlations
To gain insight into the mechanisms of intestinal dysfunction, we then analysed the association between humoral activity and clinical manifestations. The number of bowel movements per day and stool form correlated positively with jejunal mRNA expression of GLT and IGH and with the density of IgG+ cells, whereas no association was found with the intensity and frequency of abdominal pain (data not shown). The frequency of bowel movements correlated with luminal IgG and the depression score also correlated with IGH (table 3 and see online supplementary figure 3).
Correlation between clinical parameters and biological markers of humoral activity (GLT and IGH expression in jejunal mucosa, IgG in the lumen and mucosal IgG+ cells) in H and IBS-D participants
Discussion
This study demonstrates, for the first time, increased humoral activity in the jejunal mucosa of patients with IBS-D in association with cardinal clinical features of this disorder. The jejunal mucosa of these patients displays a higher density of both activated B lymphocytes and plasma cells, which locally produce increased amounts of IgG. Moreover, immunological mechanisms such as atopy, and not sex, account for differential immune activity, thereby suggesting the involvement of the mucosal-associated immune system in local immunological responses.
We identified the humoral response as one of the most significant pathways associated with the transcriptome of IBS-D, with increased production of Igs and MS4A1 (CD20 molecule), which plays a role in the development and differentiation of B cells into plasma cells. The increase in mucosal B cells is in concurs with a higher expression of gut homing integrin β7 on circulating B lymphocytes, also with increased surface IgG.14 Although previous reports did not identify changes in mucosal B cells,4 differences between our study and others may depend on the IBS subtype, intestinal region studied, method used for cell assessment and comorbid factors such as food allergy background, which correlates with the severity of IBS-D.18 Interestingly, we identified an increased proximity of plasma cells to mast cells in the mucosa of IBS-D compared with controls, which suggests a possible antibody-mediated effect of mast cells, although specific mast cell activity was not identified in the transcriptome as previously shown.1 Furthermore, NA patients displayed a greater mast cell density than controls1 ,21 and higher activation of CSR mechanisms than atopic patients. This seems contradictory because respiratory allergens also activate intestinal inflammation.34 However, additional mechanisms may account for the differences that warrant further research.
One of the major immune defence mechanisms in intestinal mucosa is the production of antibodies, of which IgA represents the main load. The loss of intestinal barrier integrity increases the chance of luminal antigens crossing the epithelium and reaching the internal milieu. Antigens entering the mucosa challenge the immune system to more specific responses such as IgG, which provides protection by eliciting inflammatory reactions.10 Indeed, IBD is associated with barrier dysfunction and high IgG mucosal production directed against intestinal bacteria,35 ,36 while IBS also shows those features, although to a lesser extent, with mucosal microinflammation and 100–1,000 times less IgG than in IBD.36 Whether mucosal IgG in IBS-D is directed against luminal components as the consequence of barrier dysfunction, and/or reflects subclinical infection, remains to be determined.
Isotype class switching favours antibody diversity, which is essential for immunological protection and takes place in organised and diffuse lymphoid tissues.37 Mucosal B lymphocytes are less activated and more scattered than those in lymphoid aggregates; however, they have been shown to maintain the capacity to class switch by mechanisms that do not involve T cells, but dendritic cells and epithelial cells.38 Bacterial antigens, through toll-like receptor (TLR) signalling via the adaptor molecule MyD88, induce the secretion of BAFF and a profileration-induced ligand (APRIL),36 cytokines that promote CSR to IgG and IgA by engaging TACI on B cells, and further cooperate in AID expression and CRS via MyD88 and nuclear factor kappa B (NF-κB).12 ,39 We showed increased IgG activity to be associated with upregulation of BAFF and APRIL receptors (BAFFR, BCMA and TACI, with a strong intercorrelation), further supported by previous demonstration by our group of increased TLR and MyD88 expression in the jejunal mucosa of IBS-D.40 Moreover, T cell-independent activation of B cells and CSR has been demonstrated by flagellin stimulation of epithelial cells cocultured with B lymphocytes,38 which could account for the increase in serum antiflagellin antibodies found in patients with IBS.13 Although not specifically addressed in our study, based on these data it seems likely that increased B cell activity in IBS-D is mediated by a T cell-independent mechanism.
The antigens responsible for eliciting such an immune response in IBS-D remain unknown; however, the role of bacterial contents as putative triggers is quite plausible because they are the main TLR pathway activators,41 and IBS is associated with altered microbiota biodiversity42 and clinical remission after antibiotic therapy.43 In our study, we only identified increased total IgG in the luminal content, and thus we do not have information on IgG subtypes being produced, and luminal IgE was below the detection limit. However, the significant increase in IGHG1, IGHG4 and IGHE mRNA points to them as the antibodies being differentially produced. Similar to intestinal IgG against microbiota components in IBD,36 the intestinal mucosa in IBS-D may also be responding to microbial proteins which mainly elicit IgG1 and, to a lesser extent, IgG3 and IgG4 production.44 Indeed, both γ1GLT and IGHG1 are highly expressed in the mucosa of IBS-D and correlate with clinical manifestations, thereby suggesting local CSR to IgG1 with clinical implications. Moreover, IgG4 could be related to IBS chronicity because prolonged antigenic stimulation with protein antigens leads to progressive expression of IgG4.44
The luminal amount of IgG depends on passive diffusion and may be the result of increased intestinal permeability in patients with IBS.19 However, neither the molecular markers of IgG production nor the protein itself was differentially identified in blood, whereas mucosal γGLT and IGHG were transcribed to a significantly higher degree in patients with IBS-D than in H subjects. It could be speculated that the differential expression of transcripts responds to differences in RNA stability. However, owing to its rapid degradation after cessation of the stimulus,45 its increased expression in the IBS-D group may be the consequence of enhanced B cell activation due to local antigenic stimulation. Moreover, the high amount of mucosal IgG+ cells in the IBS-D group points to the likelihood of luminal IgG being produced by intestinal resident plasma cells. It is noteworthy that the stress response is associated with decreased antibody production,46 more specifically the secondary immune response.47 Such a response was not observed in patients with IBS-D, who showed higher stress and depression scores than controls. By contrast, IBS-D displayed increased humoral activity, and depression positively correlated with IGHG1, IGHA and IGHG4 production. Given the importance of mucosal antibody response to homeostasis and its increase in IBS-D, additional studies are required to identify the underlying mechanisms. We wish to acknowledge the large number of variables analysed, considering the sample size and study design. In an attempt to reduce chance findings, appropriate statistical methods were applied.
In conclusion, our study revealed distinctive mucosal humoral activity associated with clinical manifestations in IBS-D. These data, together with previous observations, suggest that IBS-D does indeed have an organic basis involving mast cells, epithelial cells and T lymphocytes and also humoral mucosal activation. Identification and targeting of the antigens eliciting mucosal antibody production may be a useful approach in the management of these patients.
Acknowledgments
The authors thank M Gallart, M Casellas and C Alastrue for help in sample analysis; A Aparici, MT Casaus and P Rodríguez for invaluable assistance in the performance of jejunal biopsies; J Heredia and G Santaliestra for secretarial assistance; Christine O'Hara for editing of the manuscript and Dr A. Sanchez-Chardi and F. Cardoso from Servei de Microscòpia of Universitat Autònoma de Barcelona for excellent technical support.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
MV and AMG-C contributed equally.
Contributors MV and AMG-C designed the study, performed the experiments, analysed and interpreted the data and wrote the paper; BL, CA and MP recruited the participants, performed the biopsies and created/maintained the participants database; CM performed gene quantification analysis; MF, CS-A, ES-R and BKR-J performed analytical procedures; MG evaluated the allergy status of the participants; IdT performed the histopathological analysis; JLM performed the statistical analysis; FA, JDS and JS gave clinical advice and critically reviewed the manuscript. All authors discussed the results and commented on the manuscript at all stages.
Funding Supported in part by Fondo de Investigación Sanitaria and CIBERehd, Instituto de Salud Carlos III, Subdirección General de Investigación Sanitaria, Ministerio de Economía y Competitividad: CP10/00502 & PI13/00935 (M. Vicario); CM08/00229 (B. Lobo); CM10/00155 (M. Pigrau); FI12/00254 (ES-R); PI12/00314 (CA); EII2011-0035 (BKR-J); PI11/00716 (JS); Ministerio de Educación, Dirección General de Investigación: SAF 2009-07416 (FA); Agència de Gestió d'Ajuts Universitaris i de Recerca, de la Generalitat de Catalunya: 2009 SGR 219 (FA), 2011 BP/A00099 (CM); The Rome Foundation Award 2013 (MV); Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas: CB06/04/0021 (FA).
Competing interests None.
Patient consent Obtained.
Ethics approval Vall d´Hebron Hospital Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.