Article Text

Abstract
Objective IgG4-related disease (IgG4-RD) is a systemic disease characterised by elevated serum IgG4 and IgG4-positive lymphoplasmacytic infiltration in the affected tissues. The pathogenic role of IgGs, including IgG4, in patients with IgG4-RD, however, is unknown.
Design We examined the pathogenic activity of circulating IgGs in patients with IgG4-RD by injecting their IgGs into neonatal male Balb/c mice. Binding of patient IgGs to pancreatic tissue was also analysed in an ex vivo mouse organ culture model and in tissue samples from patients with autoimmune pancreatitis (AIP).
Results Subcutaneous injection of patient IgG, but not control IgG, resulted in pancreatic and salivary gland injuries. Pancreatic injury was also induced by injecting patient IgG1 or IgG4, with more destructive changes induced by IgG1 than by IgG4. The potent pathogenic activity of patient IgG1 was significantly inhibited by simultaneous injection of patient IgG4. Binding of patient IgG, especially IgG1 and IgG4, to pancreatic tissue was confirmed in both the mouse model and AIP tissue samples.
Conclusions IgG1 and IgG4 from patients with IgG4-RD have pathogenic activities through binding affected tissues in neonatal mice.
- CHRONIC PANCREATITIS
- IMMUNOLOGY
- PANCREATIC DISEASE
- PANCREATIC PATHOLOGY
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
IgG4-related disease (IgG4-RD) is considered an autoimmune disease due to its favourable response to steroid treatment. The pathophysiology of IgG4-RD, however, remains largely unknown.
Dramatic responses to treatment with rituximab, an anti-CD20 antibody, in patients with IgG4-RD suggest that elevated serum IgG and/or IgG4 in patients with IgG4-RD have pathogenic roles.
In pemphigus vulgaris (PV), the pathogenicity of autoantibodies was established by inducing pemphigus in neonatal mice after passive transfer of immunoglobulins from patients with PV.
What are the new findings?
Subcutaneous injection of patient IgG, but not control IgG, resulted in neonatal mouse pancreatic and salivary gland injuries.
IgG1 and IgG4 in patients with IgG4-RD have pathogenic effects on neonatal mouse pancreas and salivary gland.
This antibody in patients with IgG4-RD might recognise molecules in the extracellular matrix of neonatal mouse tissues.
How might it impact on clinical practice in the foreseeable future?
If the pathogenic effects of IgG in patients with IgG4-RD observed in this study are involved in the pathophysiology of human autoimmune pancreatitis, our data in neonatal mice might provide clues to the discovery of an autoantigen in patients with IgG4-RD.
An ELISA system detecting the autoantibodies for this antigen may be a useful tool for diagnosis and disease activity monitoring of IgG4-RD.
Introduction
IgG4-related disease (IgG4-RD) is a novel immune-mediated disorder characterised by elevated serum IgG4 levels and IgG4-positive lymphoplasmacytic infiltration in the involved organs in association with tumour-like swelling and variable degrees of tissue injury and fibrosis.1 These distinctive clinical and pathological features were first identified in patients with autoimmune pancreatitis (AIP), now referred as type 1 AIP, a unique form of chronic pancreatitis.2 ,3 Since then, simultaneous or metachronous lesions with pathological features similar to those in the pancreas have been observed in other organs,4 ,5 leading to the concept of a new systemic disease named IgG4-RD.6 ,7 Along with increased recognition of the concept of IgG4-RD, the number of the patients diagnosed with IgG4-RD is rapidly increasing. The pathophysiology of IgG4-RD, however, remains largely unknown.
Previous reports revealed the presence of several autoantibodies in the serum from patients with IgG4-RD.8–13 Moreover, it is well recognised that steroid treatment is generally very effective in patients with IgG4-RD. These facts suggest that the disease has an autoimmune nature. Recent studies demonstrated dramatic clinical responses in patients with IgG4-RD to the anti-CD20 antibody rituximab, along with significant depletion of B cells and plasmablasts.14 These findings suggest that elevated IgG and/or IgG4 levels in patients with IgG4-RD may have pathogenic roles.
IgG4 is considered a non-inflammatory, or rather anti-inflammatory, immunoglobulin because of its distinctive biological characteristics, such as the ability to exchange Fab arms,15 inability to fix complement and low affinity for Fc receptors.16 In some diseases, however, IgG4 antibodies act as tissue-destructive autoantibodies, as observed in pemphigus vulgaris (PV),17 idiopathic membranous glomerulonephritis18 and muscle-specific kinase myasthenia gravis.19 In a previous study of PV, the pathogenicity of autoantibodies was first established by inducing pemphigus in neonatal mice after passive transfer of immunoglobulins from patients with PV.17 Subsequently, IgG4 autoantibodies against intercellular adhesion molecules, desmoglein 1 and 3, were identified in patient sera as the pathogenic antibodies.20 Humans have most of the mouse genes and important domains are conserved. If human IgG has pathological effects on mice, it may have similar activities in humans.
In the present study, therefore, we injected patient IgGs into neonatal Balb/c mice and investigated the histopathological changes in the affected organs to examine the pathogenic activities of IgG and IgG subtypes derived from patients with IgG4-RD.
Materials and methods
Patients
Serum samples were obtained from 10 consecutive patients who met the comprehensive diagnostic criteria for IgG4-RD 2011,7 the diagnostic criteria of AIP21 or IgG4-related kidney disease.22 In 3 of 10 patients, additional blood samples were collected after 8 weeks treatment with prednisolone. Serum samples were also obtained from five healthy controls and five disease controls (three patients with histologically proven pancreatic cancer and two patients who met the Mayo criteria for primary sclerosing cholangitis23). Controls were age-matched (±5 years) and sex-matched with patients with IgG4-RD. An overview of the clinical characteristics of the patients and controls is provided in online supplementary table S1.
Supplementary table
Preparation of human IgG
Human IgG injected into mice was prepared from human serum samples by Ab-Rapid PuRe EX (P-015, Prote Nova, Japan) according to the manufacturer's instructions. The IgG was dialysed with phosphate-buffered saline (PBS, pH 7.2), concentrated by ultrafiltration using Amicon Ultra (UFC805024, Millipore, Germany), and stored at −20°C. Concentration of purified IgG was measured using the Human IgG EIA Kit (MK136, Takara, Japan). The IgG fraction purity was confirmed by testing for IgA, IgM, IgE and protein contamination using the Human IgA ELISA Kit (E88-102, Bethyl Laboratories, USA), Human IgM ELISA Kit (E88-100, Bethyl Laboratories), Human IgE ELISA Kit (E88-108, Bethyl Laboratories) and Coomassie Brilliant Blue staining, respectively.
Preparation of human IgG subclasses
IgG1 and IgG4 were purified from human serum samples by Capture Select IgG1 (Hu) affinity matrix (191303005, Invitrogen, USA) and Capture Select IgG4 (Hu) affinity matrix (290005, Invitrogen), respectively, according to the manufacturer's instructions. Briefly, human serum was loaded onto the column and washed with PBS. The IgG subclass fraction (IgG1 or IgG4) was eluted with 0.1 M glycine (pH 2.8) and neutralised with 1.5 M Tris (pH 7.5). The eluted IgG subclass fractions were concentrated by ultrafiltration using Amicon Ultra (UFC805024, Millipore) under extensive washing with PBS (pH 7.2). Concentrations of purified IgG subclasses were quantitated by Human IgG1 Platinum ELISA (BMS2092, eBioscience, USA), Human IgG2 Platinum ELISA (BMS2093, eBioscience), Human IgG3 Platinum ELISA (BMS2094, eBioscience) and Human IgG4 Platinum ELISA (BMS2095, eBioscience), according to the manufacturer's instructions. Subclass purity was confirmed by testing for IgA, IgM, IgE, the other IgG subclasses, and protein contamination using the protocol described above.
Concentrations of human IgGs or IgG subclasses in human and mouse sera
Concentrations of human IgG, IgG1, IgG2, IgG3 and IgG4 in human and mouse sera were measured by the methods described above.
Ex vivo mouse pancreatic organ culture assay
Ex vivo pancreatic organ culture assay was performed by following the method of the ex vivo PV assay.24 Briefly, anaesthetised 8-week-old mice (SLC, Japan) were perfused with intravenous administration of PBS. The pancreas was removed and trimmed by removing fat tissue and cut in half. Purified IgG (50 μL, 1 mg/mL) was injected into the pancreas using a 27-gauge needle. The pancreatic tissue was placed into a 24-well cell culture plate (662160, Greiner Bio-one, Germany) with 400 μL PBS containing the purified IgG (1 mg/mL) and maintained in a humidified incubator with 5% CO2 at 37°C. After 1 h culture, the pancreatic tissue was rinsed with PBS three times for 20 min each, placed into OCT compound, and frozen at −30°C.
Histological evaluation
See online supplementary methods.
Immunohistochemical study
The immunohistochemical study was performed according to standard methods for mouse and human tissue sections. The primary antibodies used were CD3 1:100 (ab16669, Abcam, England), CD45R 1:100 (ab64100, Abcam), F4/80 1:100 (ab6640, Abcam), amylase 1:100 (ab21156, Abcam), human IgG 1:100 (ab109489, Abcam) and Gr1 1:100 (14-5931-82, eBioscience), following antigen retrieval with 10 mM citrate buffer pH 6.0; and IgG4 1:100 (418051, NICHIREI, Japan), collagen IV 1:100 (ab6586, Abcam), following antigen retrieval with proteinase K. Sections were incubated with primary antibodies at room temperature for 60 min. For detection, Universal Dako LSAB+ Kit (Dako, Denmark) was used according to the manufacturer's instructions. Detection times were equally standardised for all the sections.
Immunofluorescence study
The immunofluorescence study was performed with a standard method for mouse and human tissue sections and mouse organ culture sections. For direct immunofluorescence, fluorescein isothiocyanate-conjugated rabbit antibodies specific to human IgG 1:50 (F0185, Dako), IgG1 1:50 (AF006, The Binding Site, USA), IgG2 1:50 (HP6014, Sigma-Aldrich), IgG3 1:50 (HP6050, Sigma-Aldrich) and IgG4 1:50 (HP6025, Sigma-Aldrich) were used. For indirect immunofluorescence, C1q 1:100 (ab11861, Abcam) and the same primary antibodies described in the Immunohistochemical study section were used and as secondary antibodies Alexa Fluor 488 antimouse IgG 1:100 (A-21202, Life Technologies, USA), Alexa Fluor 488 antirabbit IgG 1:100 (A-21206, Life Technologies), Alexa Fluor 594 antimouse IgG 1:100 (A-21203, Life Technologies) and Alexa Fluor 594 antirabbit IgG 1:100 (A-21207, Life Technologies) were used.
Immunoelectron microscopy study
See online supplementary methods.
Statistics
Differences were assessed using Student's t test for continuous data and the χ2 test and the Fisher's exact test for categorical data. Statistical analysis was performed using the Statistical Package for Social Sciences (SPSS V.17.0). All statistical tests were two-sided. A p value of <0.05 was considered statistically significant.
Study approval
Studies involving animals were performed with the approval of the institutional animal ethics committee. Human studies were performed according to the Declaration of Helsinki, and were approved by the Institutional Review Board of Kyoto University Hospital. All patients and controls provided written informed consent prior to inclusion in the study.
Results
Induction of pancreatic injury in mice by subcutaneous injection of IgG from patients with IgG4-RD
Subcutaneous injection of control IgG yielded a dose-dependent increase in the human IgG concentration in neonatal mouse serum (see online supplementary figure S1). Based on these data, the injection dose of patient IgG or each IgG subclass was determined to obtain the actual patient serum concentration in the mouse serum. The dose of the control IgG or IgG subclasses administered to mice was adjusted to that of each corresponding age-matched and sex-matched patient. As a result, serum human IgG concentrations in mice given patient IgG and mice given control IgG were statistically equivalent (19.4 vs 21.4 mg/mL, p=0.53). Under these conditions, subcutaneous injection of patient IgG into neonatal mice induced pancreatic injury (figure 1A) with a maximum change observed 12 h after the injection, whereas control IgG had no effect. Histologically, oedematous changes in the intralobular and interlobular spaces and around the pancreatic duct, acini necrosis, haemorrhage and infiltration of Gr1-positive polymorphonuclear leucocytes were observed in the mouse pancreas injected with patient IgG (figure 1A and see online supplementary figure S2A). No fibrous changes were detected. Quantification of the oedematous area revealed the development of oedema in all of the mice injected with patient IgG, but in none of the mice injected with control IgG (figure 1B). The levels of necrosis, haemorrhage and Gr1-positive cell infiltration were significantly greater in mice injected with patient IgG than in mice injected with control IgG (figure 1C). On the other hand, infiltration of macrophages (F4/80 positive), T cells (CD3 positive) and B cells (CD20 positive) was scarcely induced by injection with patient IgG (data not shown). Pancreatic injuries were induced by injection of patient IgG in a dose-dependent manner (figure 1D). Serum amylase levels were also significantly higher in mice given patient IgG than in mice given control IgG (1652 vs 1307 IU/L, p=0.021). The pathogenic effect of patient IgG on mice was significantly reduced when we used patient IgG obtained 8 weeks after the initiation of steroid treatment (see online supplementary figure S3). The dose of injected patient IgG obtained before and after steroid treatment was equal. These findings clearly demonstrated that IgG in the sera from patients with IgG4-RD had pathogenic effects on mouse pancreas.
Supplementary figures

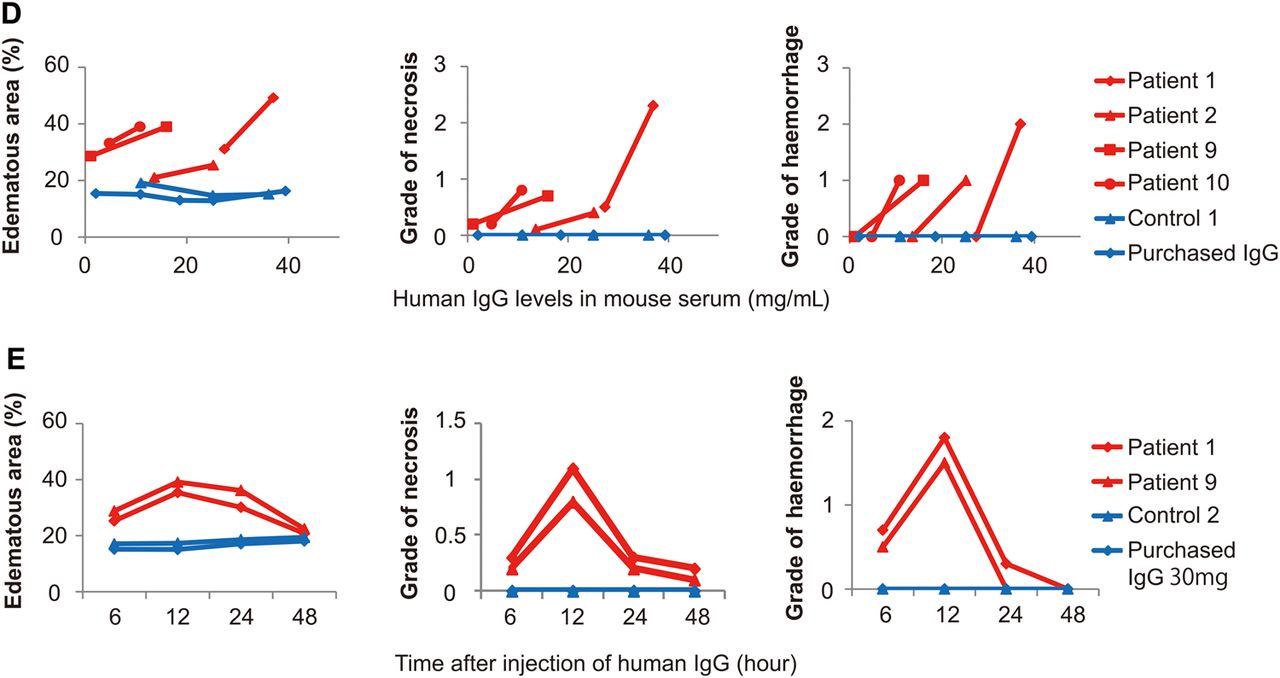
Induction of pancreatic injury in mice by subcutaneous injection of IgG from patients with IgG4-RD. Pancreatic injury 12 h after subcutaneous injection of control IgG (n=10) or patient IgG (n=10) with IgG4-RD. (A) H&E staining and immunohistochemical staining for Gr1 in pancreatic tissue sections. Scale bars: 50 μm (upper panels), 20 μm (lower panels). (B) Oedema (separation) development by IgG injection. The cut-off value of the oedematous area was defined as the mean +2 SD in mice given control IgG or PBS, 21.1%. (C) Histologic grade of necrosis, haemorrhage and Gr1-positive cell count. Data are shown as mean±SEM. *p<0.05 and **p<0.005 by paired Student's t test. (D) Dose-dependent induction of pancreatic injury in mice by injection of IgG from patients with IgG4-RD. Oedematous area, histologic grade of necrosis and haemorrhage in pancreatic tissues were assessed in mice given two different doses of IgG from the same patients (patients 1, 2, 9 and 10) or mice given several different doses of control IgG (control 1) and purchased IgG. (E) Time course of the effect of patient IgG (patients 1 and 9) or control IgG (control 2) and purchased IgG on mouse pancreas. IgG4-RD, IgG4-related disease; PBS, phosphate-buffered saline.
Induction of injuries to other organs in mice following subcutaneous injection of IgG from patients with IgG4-RD
In addition to the pancreas, we analysed salivary gland, kidney, prostate, lung, heart, liver and intestine tissue of the mice 12 h after injecting patient or control IgG. Among these organs, the salivary gland exhibited oedematous changes similar to that in the pancreas in all mice injected with patient IgG, but control IgG had no effect (see online supplementary figure S4). Unlike in the pancreas, acini necrosis, haemorrhage and infiltration of Gr1-positive polymorphonuclear leucocytes were not apparent in the salivary glands. The pathogenic activity of patient IgG on the salivary gland was also diminished when we used patient IgG obtained 8 weeks after the initiation of steroid treatment (data not shown). No pathological changes were observed in kidney, prostate, lung, heart, liver or intestine, except for oedematous changes around the lower bile duct.
Deposition of IgG and IgG subclasses from patients with IgG4-RD in mouse tissues
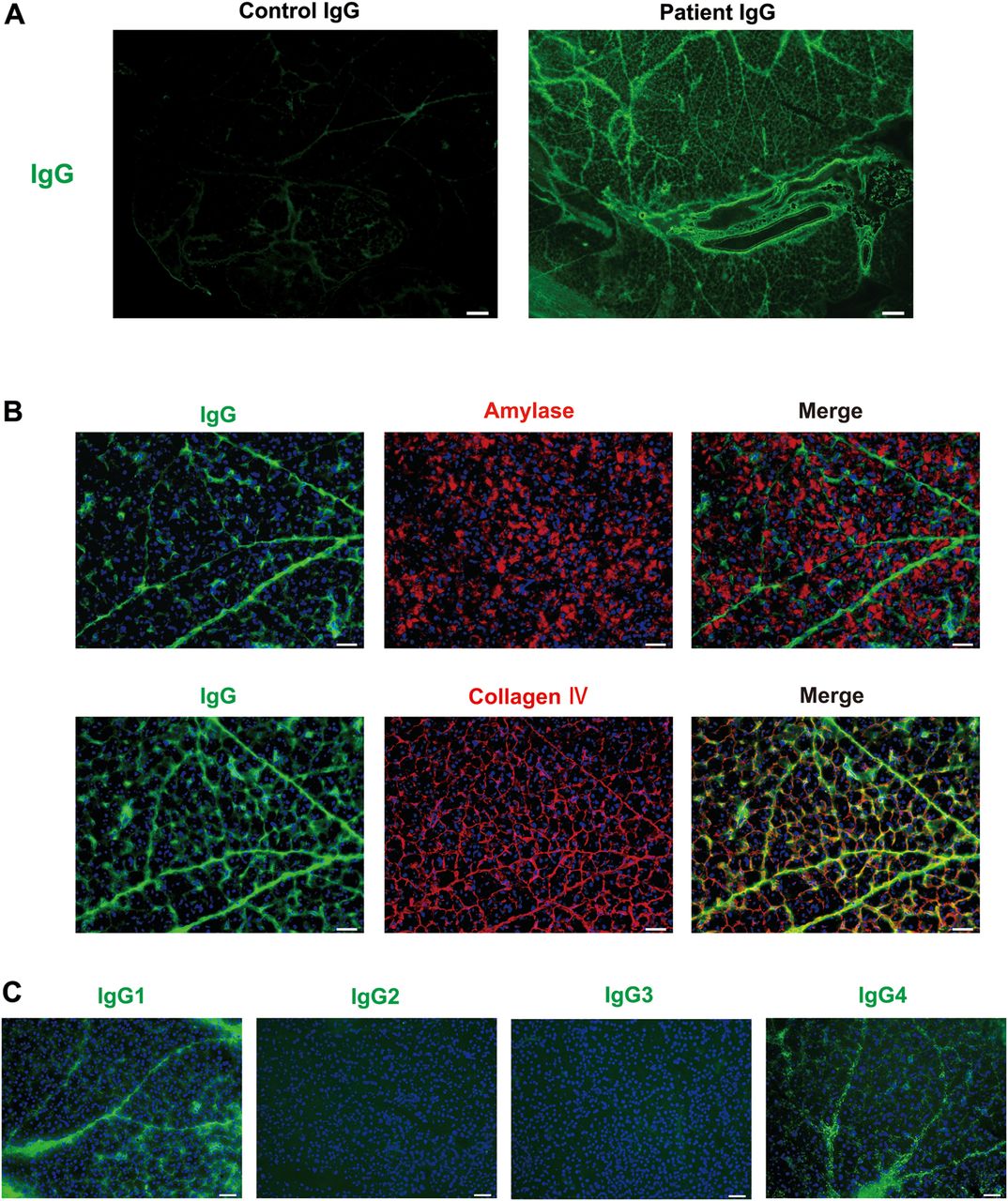
We conducted tissue-staining studies to assess whether the pathogenic activity of patient IgG is mediated by direct binding to pancreatic tissues. Immunohistochemistry detected human IgG and IgG4 deposition at the base of the acini, interlobular space (figure 2A) and around the pancreatic duct (see online supplementary figure S2B) in all mice injected with patient IgG, but not in mice injected with control IgG. IgG1 was weakly detected in 6 of 10 mice given patient IgG, but not in any mice given control IgG (figure 2A). We could not examine the deposition of IgG2 or IgG3 as antibodies for formaldehyde-fixed tissue were not available. Direct immunofluorescence also detected human IgG in pancreatic tissues in mice injected with patient IgG (figure 2B). Dual immunofluorescence studies revealed that the administered patient IgG did not colocalise with pancreatic acini expressing amylase. Most patient IgG colocalised with collagen IV expression in the basement membrane or extracellular matrix (ECM; figure 2B). The immunoelectron microscopy study revealed human IgG, IgG1 and IgG4 deposition at the base of the acini in all mice injected with patient IgG, but not in mice injected with control IgG (see online supplementary figure S5). Consistent with these results, patient IgG4 was easily visualised in the salivary glands of mice injected with patient IgG, but not in mice injected with control IgG (see online supplementary figure S6). To study the formation of immunocomplexes, we performed an immunofluorescence study to evaluate the pancreatic deposition of C1q in neonatal mice injected with patient or control IgG. Similar to human IgG deposition, C1q was stained at the base of the acini and the interlobular space in mice injected with patient IgG, but not in mice injected with control IgG (see online supplementary figure S7).

Binding of patient IgG, IgG1 and/or IgG4 to pancreatic tissues in mice injected with IgG from patients with IgG4-RD. Immunohistochemical and immunofluorescence studies 12 h after subcutaneous injection of control IgG or IgG from patients with IgG4-RD. (A) Immunohistochemical staining for IgG, IgG1 and IgG4 of mouse pancreatic tissue sections. Note that IgG and IgG4 were detected at the base of the acini and the interlobular space of pancreatic tissues in all the mice given patient IgG, but not in mice given control IgG. Scale bars: 20 μm. (B) Immunofluorescence staining of mouse pancreas. Upper panels show staining for IgG (green), amylase (red) and a merged image. Lower panels show staining for IgG (green), collagen IV (red) and a merged image. IgG was detected in collagen IV-expressing basement membrane or extracellular matrix. Scale bars: 20 μm. Representative photos are shown. IgG4-RD, IgG4-related disease.
To further analyse the binding activity of patient IgG and its subclasses to mouse pancreatic tissue, we performed an ex vivo organ culture assay using the method applied for the ex vivo PV assay.25 Patient IgG or control IgG was directly injected into pancreatic organ culture, and binding of human IgG or each IgG subclass was analysed by immunofluorescence. Similar to the in vivo study, human IgG was detected in the intralobular and interlobular spaces (figure 3A), and around the pancreatic duct (see online supplementary figure S2C), in the pancreas of mice injected with patient IgG, but not in mice injected with control IgG. Consistent with the in vivo study, as shown in figure 2B, immunofluorescence analysis in the organ culture assay also detected colocalisation of IgG with collagen IV expression, but not with amylase (figure 3B). With regard to IgG subclasses, IgG1 was detected in 6 of 10, and IgG4 in 10 of 10 pancreatic organ culture samples, whereas IgG2 and IgG3 were not or were only faintly detected (figure 3C). These data indicate that IgG, especially IgG1 and IgG4, from patients with IgG4-RD bind to mouse pancreatic tissue.
Binding of patient IgG and IgG subclasses to mice pancreatic tissues in ex vivo organ culture assay. Binding of human IgG and IgG subclasses to pancreatic tissues was assessed by immunofluorescence studies in ex vivo mice pancreatic organ cultures. (A) Immunofluorescence staining for IgG. IgG was detected in intralobular and interlobular spaces, and around the pancreatic duct in pancreatic tissues injected with patient IgG, but not in tissues injected with control IgG. Scale bars: 200 μm. (B) Upper panels show staining for IgG (green), amylase (red) and a merged image. Lower panels show staining for IgG (green), collagen IV (red) and a merged image. IgG was detected in collagen IV-expressing basement membrane or extracellular matrix. Scale bars: 20 μm. (C) Immunofluorescence staining for IgG1, IgG2, IgG3 and IgG4. IgG1 and IgG4 were detected, whereas IgG2 and IgG3 were not. Scale bars: 50 μm. Representative photos are shown.
Distinctive pathogenicity of IgG1 and IgG4 from patients with IgG4-RD
Based on the results of the mouse pancreas binding studies of each human IgG subclass (figure 3C), we next analysed the pathogenic activity of IgG1 or IgG4 from patients with IgG4-RD. IgG1 and IgG4 were prepared from patients 1, 2 and 9 and controls 1 and 2, from whom sufficient serum samples were obtained. IgG1, IgG4 alone or IgG1 + IgG4 was subcutaneously injected into neonatal mice and pancreatic injury was evaluated (figure 4). The injection dose of a patient and control IgG subclass was determined by the same method as IgG injection. Oedematous changes, acini necrosis, haemorrhage and infiltration of Gr1-positive polymorphonuclear leucocytes were induced in the pancreas of mice given patient IgG1 or IgG4, but not in mice given control IgG1 or IgG4 (figure 4A,B). Both IgG1 and IgG4 had pathogenic activity, but IgG1 had more prominent pathogenic activity than IgG4 (figure 4B). Notably, the potent pathogenic effect of patient IgG1 was significantly attenuated by simultaneous injection of patient IgG4, suggesting an inhibitory effect of IgG4 on the pathogenic activity of IgG1 (figure 4B). Indeed, immunohistochemical studies revealed stronger IgG1 staining in mice given only IgG1 than in mice given IgG1 + IgG4 (figure 4C). By contrast, the intensity of IgG4 staining in mice given only IgG4 was equal to that in mice given IgG1 + IgG4 (figure 4C). These findings suggest that IgG4 competes with IgG1 to bind to the pancreas tissue and that IgG4 has higher binding affinity than IgG1.


Pathogenic activities of IgG1 and IgG4 from patients with IgG4-RD on mouse pancreas. Pancreatic injury was evaluated in neonatal mice 12 h after subcutaneous injection of IgG1 or IgG4 alone, or IgG1 + IgG4. (A) Upper panels show H&E staining of pancreatic tissue sections of mice injected with IgG subclasses prepared from control 1. Middle panels and lower panels show H&E staining and immunohistochemical staining for Gr1, respectively, of pancreatic tissue sections in mice injected with IgG subclasses prepared from patient 9. Scale bars: 20 μm. (B) Histologic grade of necrosis, haemorrhage and Gr1-positive cell count were assessed. Data are shown as mean±SEM. *p<0.05 by paired Student's t test. Note that both IgG1 and IgG4 from patients had pathogenic activities, but IgG1 had a more pronounced effect than IgG4. This potent pathogenic activity of IgG1 was significantly diminished by simultaneous injection of the patient IgG4. (C) Immunohistochemical staining for human IgG1 and IgG4 in mouse pancreatic tissue sections injected by patient IgG1 or IgG4 or IgG1 + IgG4. Note that the intensity of IgG1 staining in mice given patient IgG1 + IgG4 was less than that in mice given only IgG1 (upper panels). In contrast, the intensity of IgG4 staining in mice given patient IgG1 + IgG4 was similar to that in mice given only IgG4 (lower panels). Scale bars: 20 μm. Similar data were obtained in all the mouse pancreatic tissue sections examined, and representative photos are shown. IgG4-RD, IgG4-related disease.
Deposition of IgG4 in the pancreatic tissue from patients with AIP
The results of the mouse studies suggested that the IgG1 and IgG4 from patients with IgG4-RD bound to the pancreatic tissue and also had some pathogenic activity (figure 4). Thus, we next performed tissue-staining studies to evaluate whether IgG1 or IgG4 was deposited in the pancreas of patients with AIP. In addition to typical infiltration of IgG4-positive plasma cells, immunohistochemistry revealed linear staining of IgG4 at the base of the acini, the interlobular space (figure 5A) and around the duct of all the patients examined (see online supplementary figure S2D). IgG1 was faintly stained in two of five patient tissues (data not shown). Immunofluorescence study also detected IgG4 in the affected areas of AIP (figure 5B,C). Similar to the results from mouse pancreas, colocalisation of IgG4 with collagen IV, but not amylase, was observed in the pancreas from patients with AIP (figure 5B), suggesting the binding of IgG4 to the basement membrane or ECM in the intralobular lesions. In the interlobular lesions, IgG4 fluorescence overlapped with collagen IV expression in the ECM (figure 5C). In contrast, normal pancreas and chronic pancreatitis tissues showed no IgG4 staining (data not shown). Immunoelectron microscopy also detected IgG and IgG1 deposition at the base of the acini in the pancreas of patients with AIP (see online supplementary figure S8), although IgG4 was not detected, probably due to technical problems.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Deposition of IgG4 to pancreatic tissue in patients with autoimmune pancreatitis (AIP). Immunohistochemical and immunofluorescence studies for pancreatic tissues in patients with AIP. (A) Upper panels show H&E staining and immunohistochemical staining for IgG4 in tissue sections from the boundary between affected (left) and non-affected (right) AIP lesions. Scale bars: 200 μm. Lower panels show high-power magnification of immunohistochemical staining for IgG4 in the areas with affected acini and interlobular space, and non-affected acini. Note that in addition to the typical staining of IgG4-positive plasma cells, linear staining of IgG4 was observed at the base of the acini and the interlobular space of the affected areas, but not in non-affected areas. Scale bars: 20 μm. (B) Immunofluorescence staining of affected acini in AIP. Upper panels show staining for IgG4 (red), amylase (green) and a merged image. Lower panels show staining for IgG4 (red), collagen IV (green) and a merged image. Note that IgG4 was bound to collagen IV-expressing basal side of the acini, merging as yellow. Scale bars: 20 μm. (C) Immunofluorescence staining of interlobular lesions in AIP showing IgG4 (red), collagen IV (green) and a merged image. IgG4 was bound to collagen IV-expressing extracellular matrix, merging as yellow. Scale bars: 20 μm. Similar data were obtained in all the patients examined, and representative photos are shown.
Discussion
In this study, by passively transferring patient IgG, we demonstrated the pathogenic effects of IgGs, including IgG4, obtained from patients with IgG4-RD on neonatal mouse pancreas. To our knowledge, this is the first report showing the pathogenicity of IgGs from patients with IgG4-RD.
According to the reported model for PV in which IgG pathogenicity was confirmed,17 ,20 we assessed the pathogenic effects of immunoglobulins from patients with IgG4-RD by injecting patient IgG into neonatal mice. Surprisingly, subcutaneous injection of patient IgG induced pancreatic lesions, whereas control IgG had no effect. Pancreatic enlargement and interlobular dissociation are the part of imaging and histopathological features of AIP, respectively.2 ,25 The oedematous lesions observed in this study may be reflected by these imaging and histological abnormalities found in patients with AIP. The haemorrhage and infiltration of polymorphonuclear cells observed in the mice are not typical findings of AIP. It should be noted, however, that polymorphonuclear cell infiltration is also detected in an animal model of PV produced by injecting mice with pathogenic IgG obtained from patients with PV, although polymorphonuclear cell infiltration is not observed in patients with PV.17 In our model, we observed no fibrotic changes, which are found in AIP, especially type 1 AIP. The reason for the absence of fibrotic changes in our mouse model is unknown at present, but a longer observation period may be required to detect the development of fibrosis. Alternatively, previous reports suggest the involvement of Foxp3 (+) T cells in the development of fibrosis in AIP.26 Indeed, we analysed neonatal mice repeatedly injected with patient or control IgG for 2 weeks after birth as a chronic model. Induction of B cell/T cell infiltration and fibrosis, however, were minimal (data not shown). In general, tissue-resident plasma cells and T cells are considered to derive from tissue-specific antigen-primed B cells and T cells.27 ,28 At this point, our model appears to lack the antigen sensitisation step for B cells and T cells, and only recapitulates the effect of the antibody on a specific tissue. This may partly explain why our model exhibited only acinar cell injury, and not the B cell/T cell responses and subsequent development of fibrosis observed in human AIP. Taken together, we speculate that the IgG in patients with IgG4-RD includes pathogenic autoantibodies against specific molecules present in the neonatal pancreas. Interestingly, Maillette de Buy Wenniger et al29 reported highly specific and dominant B cell receptor clones in the sera from patients with IgG4-RD that completely disappeared after steroid treatment. Given our findings of reduced pathogenicity of patient IgG after steroid therapy, such dominant B cell receptor clones might be responsible for the autoantibody production.
To clarify whether the pathogenic activity of the injected IgG was exerted through its direct effects on affected tissues, we assessed the binding of the injected IgG to the injured organs by immunohistochemical and immunofluorescence studies. Patient IgG, but not control IgG, was stained in the mouse pancreatic tissues. To confirm our in vivo data, we also performed ex vivo organ culture studies, as performed in the study for PV.24 Similar to the in vivo study, we found direct binding of the patient IgG to neonatal mouse pancreatic tissues. Importantly, the immunofluorescence staining of patient IgG occurred on both the basal side of the acini and the interlobular space, and overlapped with that of collagen IV, but not with amylase or cytokeratin in mouse pancreas. These results are compatible with our human data as well as with previous data presented by others showing the deposition of IgG and IgG4 at the basement membrane of the pancreatic acini and around the pancreatic duct in patients with AIP.30 ,31 Taken together, these findings suggest that the autoantibodies in patient serum may recognise molecules involved in cell–ECM adhesion. Various molecules are involved in cell–ECM adhesion in each organ. The presence of IgG autoantibodies directed at these molecules with resulting cell adhesion disruption is a common feature of several autoimmune diseases; for example, anti-BP180 antibodies in bullous pemphigoid32 and anti-BP180, laminin-332, α6 integrin or β4 integrin antibodies in mucous membrane pemphigoid.33 Notably, the predominant IgG subclasses of the autoantibodies in these diseases are IgG1 and IgG4 antibodies.
To identify the autoantigen, antigen screening was performed using mouse pancreatic tissue extracts or mouse pancreatic stellate cell extracts expressing pancreatic ECM by both immunoprecipitation experiments and western blot analysis using patient IgG and control IgG as antibodies. Some candidate proteins specific for patient IgG were identified by subsequent mass spectrometry analysis. No antibody among these candidates, including collagen IV, however, was detected by ELISA in the serum samples obtained from patients with IgG4-RD (data not shown). One possible reason for this finding is that the autoantigen was denatured or degraded by our experimental preparations or procedures. For example, the autoantigens of idiopathic membranous nephropathy (PLA2R) and Goodpasture's syndrome (type IV collagen α3) are detected in a configuration-dependent manner. Further efforts are necessary to identify the true antigen by adjusting the experimental conditions.
In the present study, we attempted to define the pathogenicity of IgG subclasses obtained from patients with IgG4-RD by passively transferring each IgG subclass into neonatal mice. Among the IgG subclasses, both IgG1 and IgG4 had binding and pathogenic activity in mouse pancreatic tissue. IgG1 was more pathogenic than IgG4 in terms of neutrophil infiltration, haemorrhage and acinar cell necrosis. Because the ability to fix complement is much weaker in IgG4 than in IgG1,16 it is reasonable to consider that the IgG1 subclass exerts direct cytotoxic activity on pancreatic tissue through the generation of immunocomplexes. Indeed, our immunofluorescence study of C1q staining strongly suggests the formation of immunocomplexes in the pancreatic tissue of mice injected with patient IgG. Consistent with this notion, decreased complement levels are often observed in patients with IgG4-RD, suggesting the involvement of complement and IgG1 immunocomplexes in the pathophysiology of IgG4-RD.34 On the other hand, in our model, we observed no significant difference in the levels of apoptosis or autophagy between the mice injected with patient IgG and those injected with control IgG (data not shown). Another important observation in this study is that patient IgG4 has dual functions. IgG4 isolated from patients with IgG4-RD induced pancreatic injury when this IgG subtype alone was injected. In contrast, IgG4 inhibited pancreatic injury induced by IgG1 from the same patients. Thus, circulating IgG4 from patients with IgG4-RD may have both pathogenic and protective roles. Importantly, the intensity of IgG1 staining in the extracellular space of the pancreas was markedly decreased when IgG1 was administered together with IgG4. In contrast, the intensity of IgG4 staining remained unchanged in the presence or absence of IgG1. Based on these data, we speculate that IgG4 recognising autoantigens in the extracellular space competes with IgG1 recognising the same antigen, and that binding of pathogenic IgG1 to the extracellular space was inhibited due to the higher binding activity of IgG4 to the extracellular space. Interestingly, a similar antagonistic function of IgG4 antibodies against pathogenic IgG1 autoantibodies was demonstrated in an experimental model of autoimmune myasthenia gravis.15
In summary, we provide new evidence of pathogenic activity of IgGs obtained from patients with IgG4-RD on neonatal mice. Whether the pathogenic effects of IgG from patients with IgG4-RD observed in this study are involved in the pathophysiology of human AIP needs to be clarified in future studies.
References
Footnotes
Contributors Study concept, design, acquisition of data, analysis and interpretation of data and drafting of the manuscript: MS, YK, TW and TC. Study supervision: TC. Statistical analysis: KY. Acquisition of data, and administrative, technical or material support: MS, TT, TMo, NK, TMa, AM, YN, TU, MT, YY, RM, YS, YO, TM AK, YS, YT, NU, KM, KN and TT. Pathological evaluation: KN and TT.
Funding This work was supported by JSPS KAKENHI 21229009, 24229005 and 24659363; the research programme of the Project for Development of Innovative Research on Cancer Therapeutics (P-Direct) from the Ministry of Education, Culture, Sports, Science and Technology of Japan; Health and Labour Sciences Research Grants for Research on Intractable Diseases from the Ministry of Health, Labour and Welfare, Japan and Practical Research Project for Rare/Intractable Diseases Grant in Japan Agency for Medical Research and Development (AMED).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Institutional Review Board of Kyoto University Hospital.
Provenance and peer review Not commissioned; externally peer reviewed.