Article Text

Abstract
Objective Barrett's metaplasia might develop if GORD causes oesophageal squamous cells to convert into columnar cells. Acid and bile exposures upregulate columnar differentiation genes like CDX2 in oesophageal squamous cells, but it is not known if such exposures downregulate squamous differentiation genes like SOX2. In addition to acid and bile, patients with GORD also have high oesophageal concentrations of nitric oxide (NO). This study aims to determine how acid, bile salts and NO affect genes that influence oesophageal cell phenotype.
Design Oesophageal squamous cells from patients with Barrett's oesophagus were exposed to acidic bile salts or NOC-9 (an NO donor). SOX2, p63 (squamous transcription factor) and CDX2 mRNAs were measured by quantitative RT-PCR. SOX2 and its regulatory Akt pathway proteins were evaluated by western blotting. S-nitrosylation by NO was blocked by dithiothreitol. Immunohistochemistry for SOX2 was performed on the oesophagus of rats with surgically induced GORD which were fed diets with and without nitrite supplementation.
Results In oesophageal squamous cells, NO profoundly decreased SOX2 protein and caused a significantly greater decrease in SOX2 mRNA than did acidic bile salts. NO also decreased p63 and increased CDX2 expression. NO caused S-nitrosylation of Akt, blocking its phosphorylation. Akt pathway inhibition by LY294002 or Akt siRNA reduced SOX2 mRNA. Rats fed with nitrite-supplemented diets exhibited weaker SOX2 oesophageal staining than rats fed with normal diets.
Conclusions In oesophageal squamous cells, NO blocks SOX2 expression through Akt S-nitrosylation. NO also increases CDX2 and decreases p63 expression. By triggering molecular events preventing squamous differentiation while promoting intestinal differentiation, NO might contribute to Barrett's pathogenesis.
- BARRETT'S METAPLASIA
- GASTROESOPHAGEAL REFLUX DISEASE
- NITRIC OXIDE
Statistics from Altmetric.com
Video abstract
Significance of this study
What is already known on this subject?
There is evidence to suggest that Barrett's oesophagus develops when mature oesophageal squamous cells or their progenitors are converted into columnar cells under the influence of GORD.
A number of studies have shown that oesophageal squamous cells exposed to acid and bile salts increase their expression of genes such as CDX2 involved in columnar cell differentiation, but little is known of how these noxious agents influence the expression of genes such as SOX2 involved in squamous cell differentiation.
In addition to having abnormal oesophageal exposure to acid and bile, patients with GORD can have high concentrations of nitric oxide (NO) in the oesophagus, but it is not clear how NO might contribute to the pathogenesis of Barrett's metaplasia.
What are the new findings?
In non-neoplastic oesophageal squamous cell lines derived from patients with Barrett's oesophagus, NO increases the expression of CDX2, decreases the expression of p63 and profoundly decreases the expression of SOX2.
NO blocks Akt activation by causing S-nitrosylation of Akt protein, and reduced Akt signalling decreases SOX2 expression.
The in vitro observation that NO downregulates the expression of SOX2 in oesophageal squamous cells is supported in a rat model of reflux oesophagitis in which animals were fed a diet supplemented with nitrite and ascorbic acid exhibit diminished staining for SOX2 in the squamous oesophagus.
How might it impact on clinical practice in the foreseeable future?
We have demonstrated that NO triggers a series of molecular events in oesophageal squamous cells that can inhibit squamous differentiation while promoting intestinal differentiation, a combination that might contribute to the pathogenesis of Barrett's metaplasia.
This study provides a rationale for research on interventions that might reduce NO production in the distal oesophagus as a way to prevent the development of Barrett's oesophagus.
Introduction
Barrett's oesophagus is the condition in which a metaplastic columnar epithelium predisposed to develop adenocarcinoma replaces oesophageal squamous epithelium that has been damaged by GORD.1 Barrett's oesophagus affects 2%–7% of adults in Western countries and is a major risk factor for oesophageal adenocarcinoma, a tumour whose rate of increase over the past four decades is among the highest reported for any cancer.2–6
The cells of origin for Barrett's oesophagus are not known. One hypothesis holds that Barrett's metaplasia develops from columnar progenitor cells in the gastric cardia that migrate into the oesophagus to replace GORD-damaged squamous cells.7 ,8 Alternative hypotheses propose that Barrett's metaplasia develops either as a result of transdifferentiation, in which mature oesophageal squamous cells convert into columnar cells, or transcommitment, in which immature progenitor cells in the oesophagus are reprogrammed to undergo aberrant columnar differentiation rather than normal squamous differentiation.9 ,10 For development of columnar metaplasia, both transdifferentiation and transcommitment involve alterations in cells that are native to the oesophagus, whereas the migration hypothesis involves cells that originate in the stomach.
Much indirect evidence suggests that transdifferentiation or transcommitment of oesophageal cells contributes to the pathogenesis of Barrett's metaplasia. Columnar epithelium lines the early embryonic oesophagus and, in a mouse model, explants of embryonic oesophagus lose columnar markers and begin to express squamous markers after 5 days in culture, with some cells expressing both columnar and squamous markers simultaneously.11 These data suggest that, in normal embryonic development, oesophageal squamous epithelium arises from columnar epithelium through a direct conversion process. By inference, a GORD-induced reversal of this process in adults could result in Barrett's oesophagus.
In patients with Barrett's oesophagus, biopsies of the junction between oesophageal squamous and metaplastic columnar epithelium can show cells with both squamous and columnar features by scanning electron microscopy,12 ,13 and light microscopy can show a peculiar multilayered epithelium with a basal layer of squamous cells covered by a superficial layer of columnar cells. It has been proposed that these changes represent transitional stages in the squamous-to-columnar metaplasia of Barrett's oesophagus.14
Further evidence for transdifferentiation or transcommitment in the pathogenesis of Barrett's metaplasia emerges from studies showing that oesophageal squamous cells and tissues exposed to gastro-oesophageal reflux in vivo or to acid and bile salts in vitro increase their expression of transcription factors involved in the development of a columnar cell phenotype such as CDX2 and the Sonic Hedgehog pathway target genes, SOX9 and FOXA2.15–20 In a rat model of surgically induced reflux oesophagitis, oesophageal squamous cells in the basal layer express Cdx genes prior to the development of intestinal metaplasia.20 Oesophageal squamous cell lines from patients with Barrett's oesophagus increase their expression of CDX2 when they are exposed to acid and bile salts,19 and CDX2 expression has been found in oesophageal squamous epithelium from patients with Barrett's oesophagus.19 ,21
The studies discussed above have focused on how GORD might contribute to development of Barrett's metaplasia by upregulating genes determining a columnar cell phenotype. However, it also seems likely that the pathogenesis of Barrett's metaplasia involves downregulation of genes such as SOX2 involved in squamous differentiation, a transcription factor that promotes development of stratified squamous epithelia.22 Mice genetically engineered to express low levels of oesophageal Sox2 develop patchy columnar lining in the oesophagus, with areas resembling human multilayered epithelium.14 ,22 Little is known of how GORD influences the expression of squamous differentiation genes such as SOX2.
Finally, studies on how GORD might underlie Barrett's pathogenesis have focused largely on effects of acid and bile salts (the major noxious components of gastric juice) on oesophageal cells. In patients with GORD, however, high oesophageal concentrations of NO can be generated from dietary nitrate in green leafy vegetables and other foods.23 This NO can react with oxygen to generate toxic, reactive nitrogen species (RNS).24 Relatively few studies have focused specifically on the role of NO in the pathogenesis of Barrett's metaplasia. Now, we have explored mechanisms whereby acid, bile salts and NO might influence the expression of SOX2 in non-neoplastic oesophageal squamous cell lines and have correlated our in vitro findings with in vivo data from a rat model of reflux oesophagitis. We have found that NO triggers molecular events in oesophageal squamous cells that can inhibit squamous differentiation while promoting intestinal differentiation, a combination that might lead to development of Barrett's oesophagus.
Materials and methods
Cell lines
We used two non-neoplastic, telomerase-immortalised oesophageal squamous cell lines (normal oesophageal squamous (NES)-B3T, NES-B10T, created from endoscopic biopsy specimens of the distal, squamous-lined oesophagus of two patients with Barrett's oesophagus) developed in our laboratory.19 ,25 All cell lines were maintained at 37°C in a 5% CO2 incubator.
Acidic bile salt and NO exposure
For individual experiments, NES cells were cultured in neutral medium (pH 7.2) alone; neutral medium with periodic exposures to 500 µM of 6-(2-Hydroxy-1-methyl-2-nitrosohydrazino)-N-methyl-1-hexanamine (NOC-9; Sigma, St Louis, Missouri, USA), which is a small molecule NO donor; or neutral medium with periodic exposures to acidic medium (pH 5.5) containing a mixture of conjugated bile acids as previously described.26 NO measurements were performed using the ISO-NO Mark II NO metre and ISO-NO sensor (WPI, Florida, USA) as previously described.27
Semiquantitative and quantitative real-time PCR (qRT-PCR)
Semiquantitative and qRT-PCR were performed as previously described.28 The primer sequences (table 1) were designed using Primer Express (Applied BioSystems, Foster City, California, USA) and manufactured by Integrated DNA Technologies (Coralville, Iowa, USA).
Oligonucleotide primers
Protein extraction and immunoblotting
Protein extracts were subjected to immunoblot analyses using primary antibodies (see online supplementary table S1) overnight at 4°C. As a loading control, membranes were stripped and re-probed with mouse anti-β-tubulin (Sigma, St. Louis, Missouri, USA).
Inhibition of Akt signalling
Inhibition of Akt signalling was performed using the highly selective pharmacological inhibitor LY294002 (Cell Signaling, Danvers, Massachusetts, USA)29 or a specific Akt siRNA (Thermo Scientific, Waltham, Massachusetts, USA) per manufacturer's instructions.
Mechanistic target of rapamycin-Rictor kinase assay
To determine whether NOC-9 might reduce Akt phosphorylation by decreasing rictor-mechanistic target of rapamycin (mTOR) activity, we used an in vitro kinase assay.30 Following immunoprecipitation of rictor, exogenous inactive Akt/PKB1 protein (that could be phosphorylated at Ser473 by the immunoprecipitated rictor) was added and western blots performed for phospho-Akt Ser473 as an indicator of rictor-mTOR activity.
S-nitrosylation of Akt
During S-nitrosylation, NO converts protein cysteine thiols into S-nitrosothiols, which can be detected using a biotin-switch assay. S-nitrosylation can also be blocked by the reducing agent, dithiothreitol (DTT; Invitrogen, Carlsbad, California, USA).31
Immunohistochemistry in a rat model of reflux oesophagitis
For immunohistochemical studies, we used previously collected oesophageal squamous tissue sections from rats with surgically induced reflux oesophagitis which were fed diets with and without NO supplementation as described.32 Tissue sections were obtained 4 weeks after the operation from animals with (n=6) or without (n=8) dietary NO supplementation.32 At this time point, the six animals given dietary NO supplementation developed intestinal metaplasia, while the eight animals fed diets without NO supplementation did not.32 Tissues were incubated with 1:100 dilution of SOX2 (Abcam, Cambridge, Massachusetts, USA) and target protein was visualised using diaminobenzidine as substrate. Tissues were evaluated and scored by two GI pathologists (AA, RDO) blinded to the treatment groups.
Data analyses
Quantitative data are expressed as mean±SEM. Statistical analyses were performed using an unpaired Student's t test or, for multiple comparisons, an analysis of variance and the Student-Newman-Keuls multiple-comparisons test was used. p Values ≤0.05 were considered significant for all analyses (additional details can be found in the online supplementary Material and methods section).
Results
NOC-9 treatment profoundly suppresses SOX2 mRNA and protein expression in oesophageal squamous cells from patients with Barrett's oesophagus
Exposure to acidic bile salts significantly reduced SOX2 mRNA expression in NES-B10T cells, but not NES-B3T cells (figure 1A). In contrast, exposure to NOC-9 decreased SOX2 mRNA expression in both cell lines to levels significantly lower than in untreated control cells and in cells treated with acidic bile salts (figure 1A). NOC-9 also decreased SOX2 protein levels in both oesophageal squamous cell lines (figure 1B). After reviewing these data, we elected to use NOC-9 (rather than acidic bile salts) for all further experiments, given NOC-9's consistent and profound suppression of SOX2 mRNA and protein.

Treatment with NOC-9 suppresses SOX2 mRNA and protein expression in oesophageal squamous cell lines (normal oesophageal squamous (NES)-B3T and NES-B10T) from patients with Barrett's oesophagus. Figures show representative experiments of (A) qRT-PCR for SOX2 mRNA and (B) western blotting for SOX2 protein in NES cell lines following 5 days of treatment for 1 h, 3 times daily with either acidic bile salts (A and B) or 500 µM NOC-9; mRNA or protein was collected on the following day (day 6). Bar graphs represent the mean±SEM. ***p≤0.001 compared with non-treated control; +++p≤0.001 compared with acidic bile salt-treated cells. (C) Continuous measurements of nitric oxide (NO) concentrations generated from media containing NOC-9 in doses of 250, 500 and 1000 µM.
We measured NO concentrations generated from media containing NOC-9 and found that NOC-9 doses of 250, 500 and 1000 µM generated 15, 30 and 60 µM peak concentrations of NO, respectively (figure 1C). Such concentrations have been detected in the oesophageal lumen of patients with GORD and Barrett's oesophagus following nitrate ingestion.33 Concentrations of NO peaked within 2 min after NOC-9 exposure (figure 1C).
NOC-9-mediated decrease in SOX2 is associated with decreased expression of mRNAs for TA and ΔN isoforms of p63, and increased expression of CDX2 mRNA
p63 is a transcription factor required for development and maintenance of oesophageal stratified squamous epithelium.22 ,34 The p63 gene encodes two major protein isoforms, TAp63 and ΔNp63, and p63 gene expression is normally linked with that of SOX2 in the oesophagus. To determine whether NOC-9 decreases p63 as well as SOX2, we determined mRNA expression for p63 isoforms using qRT-PCR. Treatment with NOC-9 significantly decreased both TAp63 and ΔNp63 mRNAs in NES-B3T and NES-B10T cells (figure 2A, B). CDX2, a transcription factor that determines intestinal development, appears to have an inverse relationship with SOX2.35 ,36 Accordingly, NOC-9 treatment that caused a decrease in SOX2 caused an increase in CDX2 mRNA expression in both NES-B3T and NES-B10T cells, but the increase reached statistical significance only in NES-B3T cells (figure 2C).

NOC-9 suppresses mRNA expression of TA and ΔN isoforms of p63 and increases mRNA expression of CDX2 in normal oesophageal squamous (NES) cells. Figures show representative experiments for qRT-PCR of (A) TAp63, (B) ΔNp63, and (C) CDX2 mRNA expression in NES cell lines following 5 days of treatment with 500 µM NOC-9 for 1 h, 3 times daily; mRNAs were collected on the following day (day 6). Bar graphs represent the mean±SEM. *p≤0.05 compared with non-treated control; ***p≤0.001 compared with non-treated control.
In a rat model of surgically induced reflux oesophagitis, an NO-supplemented diet decreases SOX2 expression in the distal squamous oesophagus
In a rat model, oesophagojejunostomy and gastrojejunostomy were performed to induce gastro-duodenal-oesophageal reflux, and the rats developed reflux oesophagitis and columnar metaplasia similar to Barrett's oesophagus.32 The development of metaplasia was hastened by an NO-supplemented diet. To determine NO effects on oesophageal Sox2 expression in this model, we performed immunohistochemistry for Sox2 on the distal squamous-lined oesophagus of rats that were fed postoperative diets with and without NO supplementation.32 We found Sox2 staining predominantly in the nuclei of the basal and suprabasal cells of squamous epithelium in all animals (figure 3). However, the intensity of Sox2 staining was weaker in rats fed with an NO-supplemented diet (0.83±0.17 SEM) than in rats fed with a normal diet (1.25±0.16) (figure 3).

In a rat model of reflux oesophagitis, dietary nitric oxide (NO) supplementation reduces Sox2 expression in the distal squamous oesophagus. Representative photomicrographs of the distal squamous oesophagus of rats with surgically induced reflux oesophagitis that were fed diets with (lower panels) or without (upper panels) NO supplementation at 4 weeks after surgery. Inflammatory changes are seen by H&E staining in both groups. Sox2 staining is seen predominantly in the nuclei of basal and suprabasal cells, and the intensity of Sox2 staining is weaker in the rats fed with an NO-supplemented diet.
NOC-9 does not cause increased BMP signalling
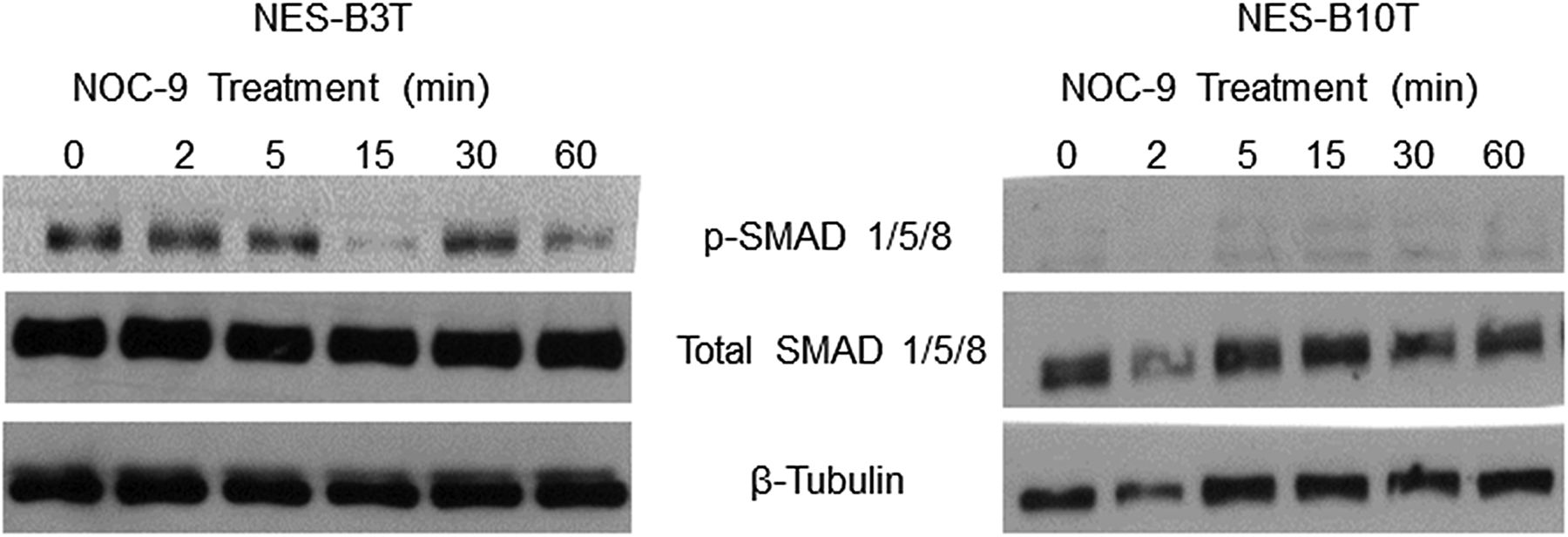
In non-oesophageal cells, activation of bone morphogenetic protein (BMP) pathway signalling has been linked with decreased SOX2 expression.35 ,37 Therefore, we investigated whether NOC-9 might decrease SOX2 expression in NES cells by activating BMP signalling. We performed western blots for the phosphorylated form of SMAD1/5/8, an indicator of BMP pathway signalling. We found no increase in p-SMAD1/5/8 in either NES-B3T or NES-B10T cells following NOC-9 treatment (figure 4). This suggests that NOC-9 does not decrease SOX2 expression by increasing BMP pathway signalling in oesophageal squamous cells.
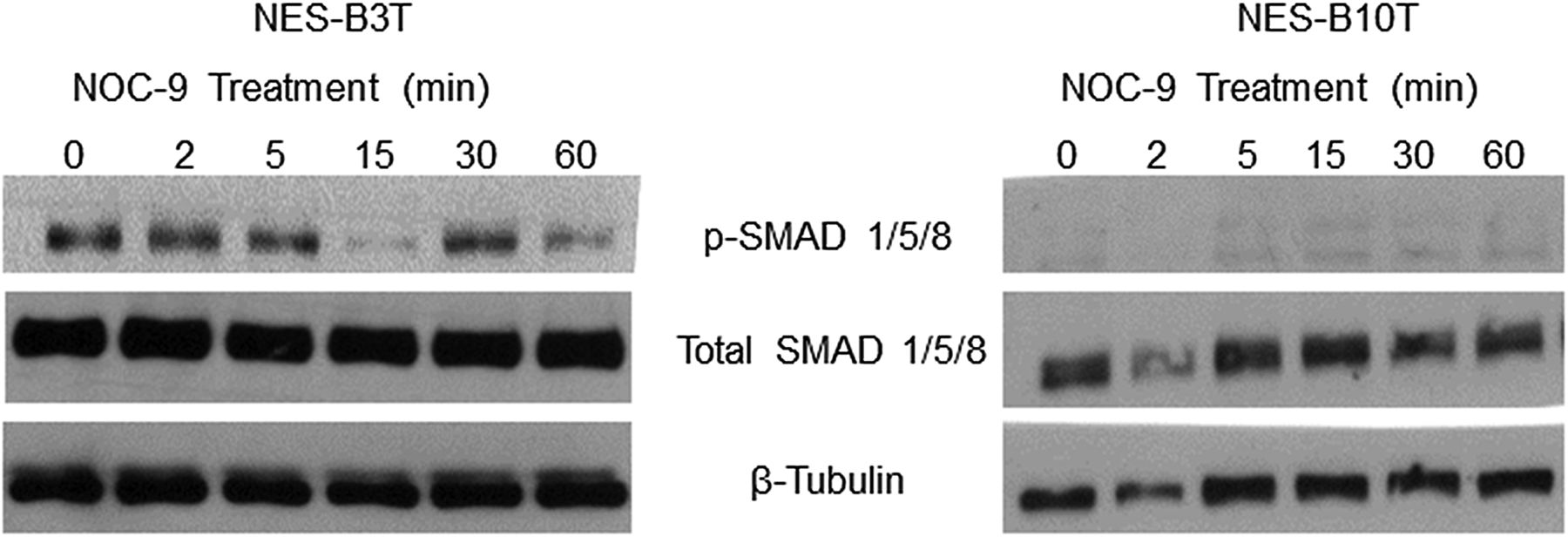
NOC-9 treatment does not increase bone morphogenetic protein (BMP) pathway signalling in normal oesophageal squamous (NES) cells. Representative experiments of western blotting for phosphorylated and total SMAD 1/5/8. NOC-9 (500 µM) was administered at time 0, and the results show the effects at the indicated time points after NOC-9 administration.
NOC-9 decreases phospho-Akt expression, and Akt signalling regulates SOX2 expression
Suppression of Akt signalling has been linked with decreased expression of SOX2 in non-oesophageal cells.38 ,39 For full activation, Akt must be phosphorylated at two distinct sites, that is, threonine 308 (Thr308) and serine 473 (Ser473).40 We treated NES cells with NOC-9 and determined the effects on Akt phosphorylation at both sites. In both cell lines, NOC-9 decreased phospho-Akt at Ser473 and at Thr308, at 2 and 5 min after exposure (figure 5A). To confirm that Akt signalling was indeed linked with SOX2 expression in oesophageal squamous cells, we inhibited Akt using the pharmacological inhibitor LY294002 or a specific siRNA, and assessed SOX2 expression by qRT-PCR. Semiquantitative RT-PCR was performed to determine the efficiency of siRNA for inhibiting Akt expression. LY294002 significantly decreased SOX2 expression in NES-B10T cells (figure 5B), a finding that was confirmed using siRNA against Akt in both NES-B3T and NES-B10T cells (figure 5C). These data demonstrate that NOC-9 decreases phosphorylation of Akt at both Thr308 and Ser473, and that Akt signalling mediates expression of SOX2 in oesophageal squamous cells from patients with Barrett's oesophagus.
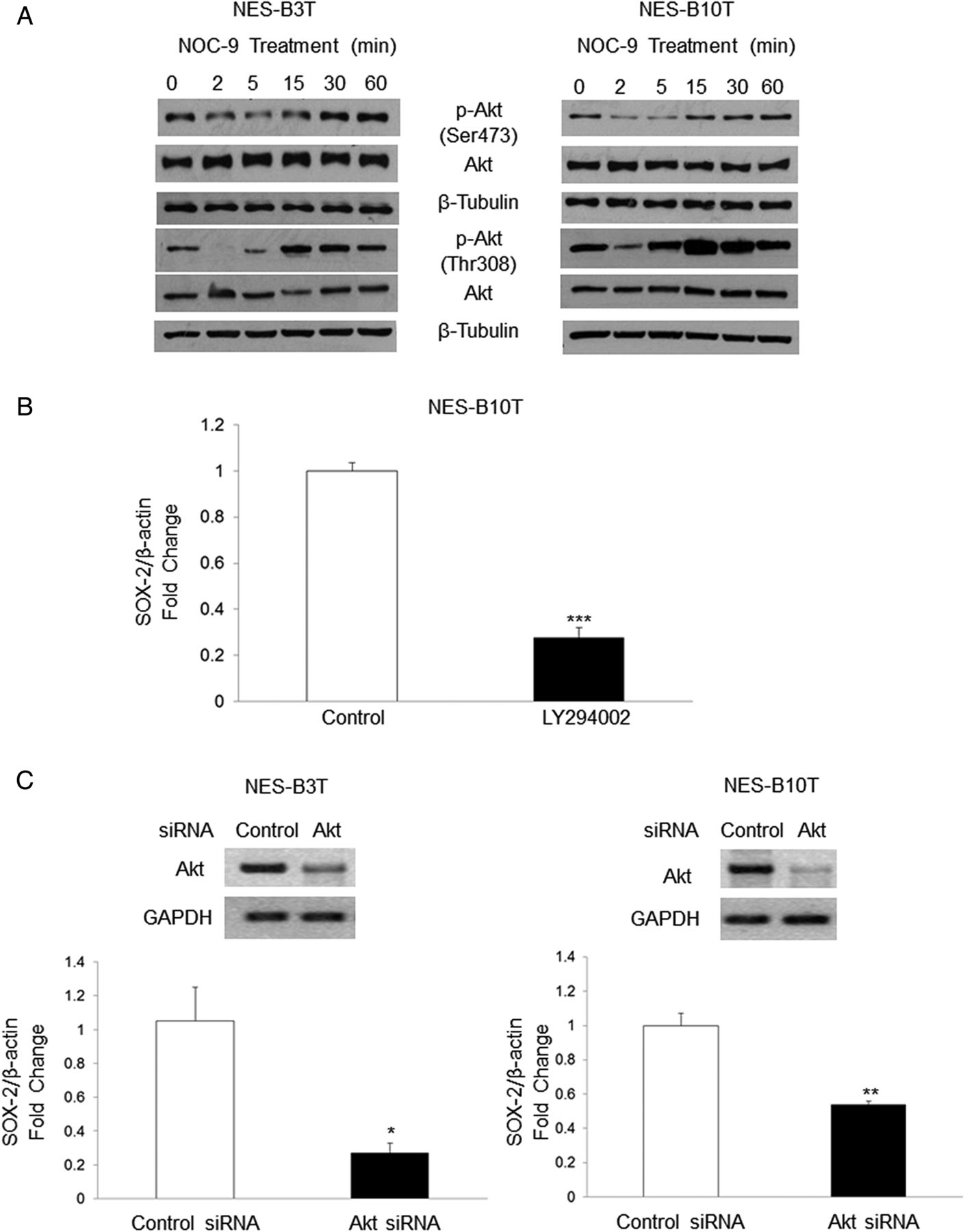
Treatment with NOC-9 decreases phosphorylation of Akt at Ser473 and Thr308, and inhibition of Akt signalling decreases SOX2 mRNA in normal oesophageal squamous (NES) cells. (A) Representative experiments of western blotting for phospho-Akt Ser473 and Thr308. NOC-9 (500 µM) was administered at time 0, and the results show the effects at the indicated time points after NOC-9 administration. Representative experiment of qRT-PCR for SOX2 mRNA in NES cells treated with (B) LY294002 for 24 h or (C) a specific Akt siRNA for 48 h; gels demonstrate semiquantitative RT-PCR showing Akt knockdown by siRNA at 48 h. Bar graphs represent the mean±SEM. *p≤0.05 compared with non-treated control; **p≤0.01 compared with non-treated control; ***p≤0.001 compared with non-treated control.
NOC-9 reduces Akt phosphorylation rapidly without affecting PI3 kinase or rictor-mTOR activity
PI3 kinase activity initiates Akt phosphorylation by recruiting Akt to the plasma membrane, along with PDK1 (3-phosphoinositide-dependent kinase-1), which phosphorylates Akt at Thr308.30 ,40 We explored whether NOC-9 might decrease Akt phosphorylation by inhibiting activation (phosphorylation) of PI3 kinase and found that NOC-9 caused no apparent decrease in PI3 kinase phosphorylation in either cell line (figure 6A).

Early NOC-9-induced decreases in Akt phosphorylation are independent of effects on PI3 kinase or rictor-mechanistic target of rapamycin (mTOR) complex activity in normal oesophageal squamous (NES) cells. (A) Representative experiments of western blotting for phosphorylated and total PI3K; *non-specific bands (B) Representative experiments of western blotting for total and phosphorylated-Akt Ser473 in a rictor-mTOR activity assay (upper blots) or in whole cell lysates (lower blots) from cells treated with 500 µM NOC-9 or (C) NES-B10T cells treated with 1000 µM NOC-9. NOC-9 (500 µM or 1000 µM) was administered at time 0, and the results show the effects at the indicated time points after NOC-9 administration.
Phosphorylation of Akt at Ser473 is accomplished by an mTOR complex that contains rictor (rapamycin-insensitive companion of mTOR).30 To determine whether NOC-9 might reduce Akt phosphorylation by decreasing rictor-mTOR activity, we used an in vitro kinase assay in which we immunoprecipitated rictor from NES cells treated with NOC-9, then added exogenous inactive Akt/PKB1 protein (that could be phosphorylated at Ser473 by the immunoprecipitated rictor) and performed western blots for phospho-Akt Ser473 as an indicator of rictor-mTOR activity. We also compared the time course of NOC-9 effects on phosphorylation of exogenous Akt by immunoprecipitated rictor with those on phosphorylation of endogenous Akt in whole cell lysates. NOC-9 had little effect on phosphorylation of exogenous Akt at Ser473 by immunoprecipitated rictor at any time point (figure 6B; upper blots). In contrast, a profound decrease in phosphorylation of endogenous Akt at Ser473 was apparent in the whole cell lysates after only 2 min of NOC-9 treatment (lower blots). These effects were even more apparent when the cells were treated with a higher concentration of NOC-9 (1000 µM rather than 500 µM, figure 6C). This high-concentration NOC-9 virtually abolished Akt phosphorylation at Ser473 within 1 min, whereas NOC-9 effects on immunoprecipitated rictor were not apparent until much later. These findings demonstrate that the rapid effect of NOC-9 in reducing Akt phosphorylation in NES cells is independent of the effects on PI3 kinase or on rictor-mTOR activity.
NOC-9 cause S-nitrosylation of Akt
It is known that NO can inactivate Akt protein through S-nitrosylation in which NO converts cysteine thiols into S-nitrosothiols.31 ,41 This S-nitrosylation can be blocked by the reducing agent DTT.41 We treated NES-B3T and NES-B10T cells with NOC-9 in the presence or absence of DTT and found that DTT attenuated or abolished the reduction in phospho-Akt Ser473 (figure 7A, B, top panels) and Thr308 (figure 7A, B, bottom panels) at 2 and 5 min. To confirm that these findings were the result of S-nitrosylation, we performed biotin-switch assays in NES-B10T cells treated with NOC-9. We found increased levels of biotinylated Akt in cells treated with NOC-9 in the presence of ascorbate (confirming S-nitrosylation) at 2 and 5 min after exposure (figure 7C). These findings show that NO causes S-nitrosylation of Akt, which interferes with its phosphorylation at Ser473 and Thr308. A schematic model summarising proposed mechanisms based on our findings is provided in figure 8.
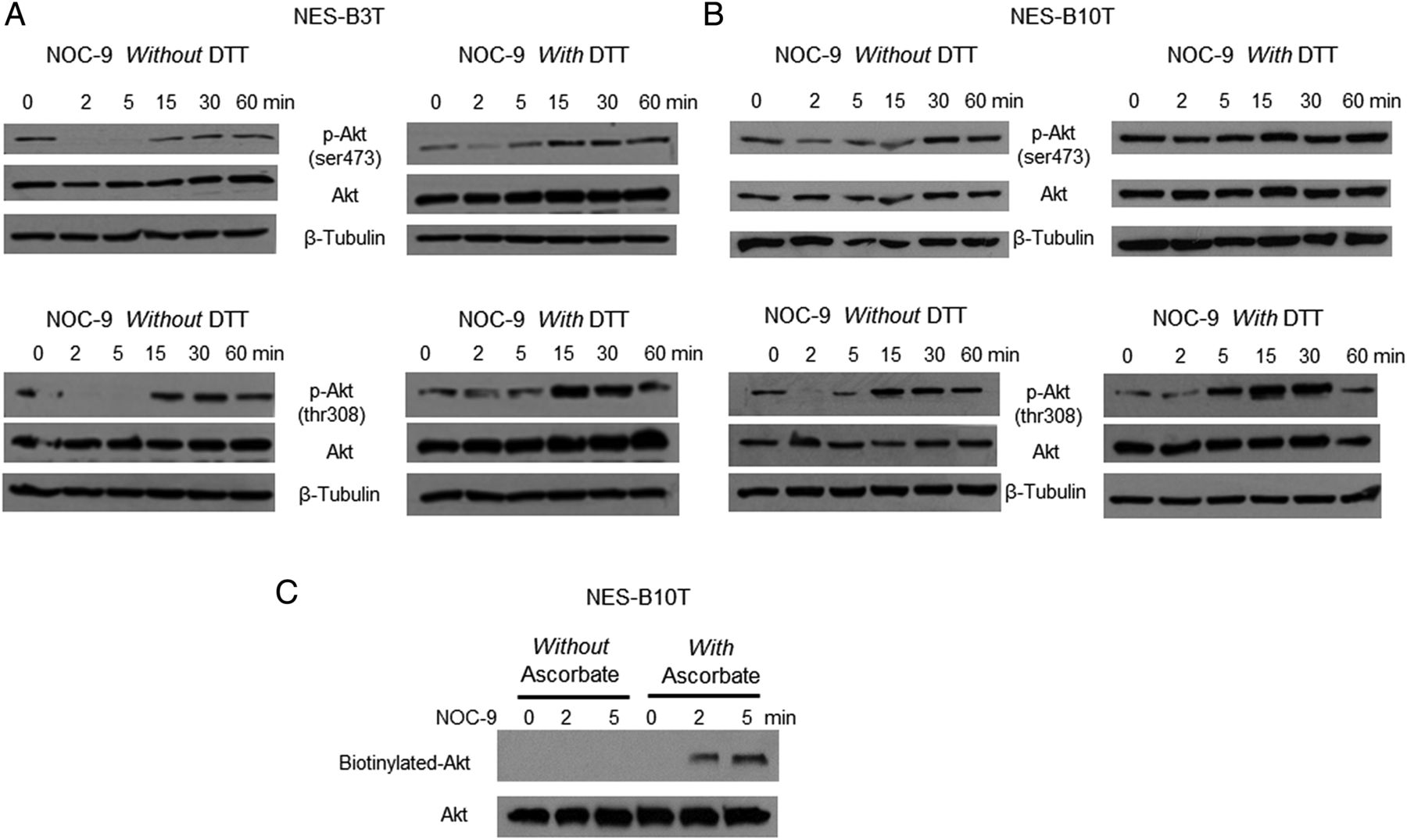
Treatment with NOC-9 induces Akt S-nitrosylation, which interferes with its phosphorylation in normal oesophageal squamous (NES) cells. Representative experiments of western blotting for total and phospho-Akt Ser473 and Thr308 with and without dithiothreitol (DTT) in (A) NES-B3T and (B) NES-B10 T cells. (C) Representative biotin-switch assays in NES-B10T cells. Note the appearance of biotinylated Akt in cells treated with ascorbate, confirming S-nitrosylation. NOC-9 (500 µM) was administered at time 0, and the results show the effects at the indicated time points after NOC-9 administration.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Schematic model demonstrating the effects of nitric oxide (NO) on Akt, SOX2, p63 (dashed line) and CDX2 (dotted line) in oesophageal squamous cells. NO causes Akt S-nitrosylation (SNO), which blocks its activation by PI3K and rictor-mechanistic target of rapamycin (mTOR), leading to decreased expression of SOX2 mRNA and protein. SOX2 is known to promote the development of stratified squamous epithelium. NO also decreases the expression of p63 isoforms, which also are required for the development of stratified squamous epithelium. At the same time, NO increases the expression of CDX2, which promotes intestinal differentiation. Thus, NO triggers molecular events that inhibit squamous differentiation while promoting intestinal differentiation, a combination that might lead to the development of the intestinal metaplasia of Barrett's oesophagus.
Discussion
In oesophageal squamous epithelial cells from patients with Barrett's oesophagus, we have demonstrated that SOX2 mRNA expression is suppressed by exposure to acidic bile salts or NO in physiologically relevant concentrations. However, these transcriptional suppressive effects are significantly greater with NO than with acidic bile salts, and NO also profoundly decreases expression of SOX2 protein. We have shown that NO causes S-nitrosylation of Akt protein, which blocks its phosphorylation at both Thr308 and Ser473, and that interference with Akt signalling in oesophageal squamous cells reduces their expression of SOX2. We have also shown that the squamous-lined distal oesophagus of rats with surgically induced reflux oesophagitis exhibits diminished staining for SOX2 when the animals are fed a diet supplemented with NO. Finally, we have demonstrated that NO decreases mRNA expression of p63 isoforms, while increasing mRNA expression of CDX2 in oesophageal squamous cells. Thus, NO triggers molecular events that can inhibit squamous differentiation while promoting intestinal differentiation in the oesophagus. These observations elucidate molecular mechanisms whereby NO generated in the oesophagus from dietary nitrate might contribute to the development of Barrett's metaplasia.
In earlier reports, we proposed that differences among individuals in oesophageal molecular pathways activated by GORD might determine whether that GORD is complicated by development of Barrett's metaplasia.16 ,19 This proposal emerged from our observations that oesophageal squamous cells from patients with and without Barrett's oesophagus responded differently to noxious stimuli. For example, acidic bile salts increased CDX2 expression in oesophageal squamous cells from patients with Barrett's oesophagus, but not in those from patients who had GORD without Barrett's oesophagus.19 We also observed differences between oesophageal squamous cells from patients with and without Barrett's oesophagus in signalling through Hedgehog, a key developmental pathway known to influence genes that determine an intestinal phenotype.16 Based on these observations, we elected to use oesophageal squamous cell lines from patients with Barrett's oesophagus for the studies described in this report.
Most investigations on how noxious agents in refluxed gastric juice might contribute to development of Barrett's metaplasia have focused on how those agents increase expression of transcription factors that determine a columnar cell phenotype such as CDX2. However, it seems equally plausible that the pathogenesis of a squamous-to-columnar metaplasia should also involve decreased expression of transcription factors that determine a squamous cell phenotype, and few studies have addressed this issue. We have focused on how noxious agents affect SOX2, an endodermal transcription factor that plays a key role in formation of stratified squamous epithelium in the embryo.22 In mice genetically engineered to express oesophageal Sox2 at levels <50% of normal, the embryonic oesophagus develops regions of columnar lining with foci of multilayered epithelium similar to those described in Barrett's oesophagus, and the oesophagus expresses genes normally expressed in the glandular stomach (eg, trefoil factors 1 and 2, Muc5B).22 ,42 Thus, genetically induced SOX2 reductions in the embryonic mouse oesophagus shift its gene expression patterns and histological features towards a columnar phenotype. Decreases in oesophageal SOX2 expression have been found in the setting of reflux oesophagitis in animal models and in patients with Barrett's oesophagus.14 However, very little was known about the mechanisms whereby reflux might downregulate SOX2.
To study how noxious agents in refluxed gastric juice affect oesophageal expression of genes influencing cell phenotype, we treated oesophageal squamous cells with acidic bile salts or with the NO donor NOC-9, using concentrations and exposure durations designed to simulate typical reflux episodes.23 ,43–46 Although most studies on this issue have focused primarily on effects of acid and bile salts, NO in the oesophagus can react with oxygen to form RNS that can have considerable toxic effects.24 NO is generated from nitrate (NO3−) ingested largely in green leafy vegetables. Some of this nitrate is concentrated in the salivary glands and secreted into the mouth, where bacteria reduce the recycled nitrate to nitrite (NO2−). When swallowed, nitrite encounters acidic gastric juice containing ascorbic acid, a powerful reducing agent, and nitrite is converted rapidly to NO. In a study of healthy volunteers who ingested nitrate in an amount found in typical salad portions, potentially genotoxic concentrations of NO (up to 50 µM) were generated at the gastro-oesophageal junction, the location where nitrite in swallowed saliva normally encounters acidic gastric juice.23 In patients with GORD and Barrett's oesophagus, who frequently reflux acid into the oesophagus, high oesophageal concentrations of NO (up to 60 µM) are generated after nitrate ingestion.33 We found that both acidic bile salts and NO suppressed SOX2 expression in oesophageal squamous cells, but SOX2 suppression induced by NO was significantly greater than that for acidic bile salts.
The transcription factor p63, a homologue of the tumour suppressor gene p53, is required for development and maintenance of oesophageal stratified squamous epithelium.22 ,34 ,47 Embryonic p63-knockout mice develop an oesophagus lined by columnar cells.48 The p63 gene encodes two major, functionally distinctive protein isoforms, TAp63 and ΔNp63. Studies in squamous carcinoma cell lines have demonstrated that SOX2 and ΔNp63 proteins interact physically and cooperate functionally to coregulate scores of downstream target genes.34 We found that exposure to NO, in addition to reducing SOX2 expression, also decreased mRNA expression of both the TA and ΔN isoforms of p63 in oesophageal squamous cells.
SOX2 and CDX2, transcription factors that determine an intestinal-type columnar cell phenotype, appear to have an inverse relationship.35 ,36 Downregulation of SOX2 and upregulation of CDX2 have been associated with intestinal metaplasia in the stomach.35 ,49 ,50 Further supporting an inverse relationship for these transcription factors, we found that exposure to NO significantly decreased SOX2 and increased CDX2 mRNA expression in our oesophageal squamous cell lines. We also analysed tissue specimens of squamous-lined distal oesophagus in rats with surgically induced GORD that were fed diets with or without NO supplementation. We found weaker nuclear staining for SOX2 in rats fed with an NO-supplemented diet than in those on normal diets. Furthermore, at 4 weeks after surgery, rats fed with NO-supplemented diets developed intestinal metaplasia with Cdx2 expression significantly more often than rats fed with normal diets.32
In certain non-oesophageal cells, decreases in SOX2 expression have been linked either with activation of BMP pathway signalling or with suppression of Akt signalling.35 ,38 ,39 We found that NO did not cause appreciable activation of BMP signalling, but did suppress Akt activation markedly. Akt is a serine/threonine kinase with critical regulatory roles in diverse cellular processes including cell differentiation.51 Recent studies have shown that Akt signalling is a key regulator of SOX2 expression and function.41 ,52 In keeping with these reports, we have found that inhibition of Akt signalling using either a pharmacological inhibitor or a specific siRNA decreases SOX2 mRNA expression in oesophageal squamous cells.
Akt is activated by phosphorylation both at Thr308 (located in the protein's activation T-loop) and at Ser473 (located in the hydrophobic motif of the protein's C-terminal tail).30 ,40 Phosphorylation at Thr308 is initiated by PI3K, which recruits Akt and PDK1 to the plasma membrane, where PDK1 phosphorylates Akt at Thr308.40 Akt phosphorylation at Ser473 is accomplished through kinase activity of the rictor-mTOR complex, an event that might also depend on PI3K activity.30 ,40 We have found that NO disrupts Akt signalling in oesophageal squamous cells by causing S-nitrosylation of Akt protein. S-nitrosylation is the covalent binding of NO to protein cysteine residues, forming S-nitrosothiols that can interfere with protein function through effects on protein conformation.53 ,54 S-nitrosylation of Akt, which disrupts its kinase activity, can be blocked by the reducing agent DTT.31 ,41 We found that DTT blocks the NOC-9-induced decrease in phosphorylation of Akt at Thr308 and Ser473, indicating that S-nitrosylation of Akt interferes with its phosphorylation. Conceivably, S-nitrosylation of PI3K could interfere with Akt signalling, but we found that NOC-9 treatment caused no reduction in phosphorylated PI3K in oesophageal squamous cells. Using an in vitro kinase assay, we also found that NOC-9 had no appreciable early effect on the ability of rictor-mTOR to phosphorylate exogenous Akt at Ser473. Furthermore, NOC-9 in high concentration virtually abolished phosphorylation of endogenous Akt at Ser473 within 2 min, a time point at which NOC-9 did not affect the ability of rictor-mTOR to phosphorylate exogenous Akt at Ser473. These observations demonstrate that NO does not reduce Akt phosphorylation by decreasing PI3K or rictor-mTOR activity and further support Akt protein S-nitrosylation as the mechanism whereby NO disrupts Akt phosphorylation and signalling.
In conclusion, in oesophageal squamous cells from patients with Barrett's oesophagus, we have shown that NO causes Akt S-nitrosylation, which blocks its activation by PI3K and rictor-mTOR, and we have shown that NO decreases expression of SOX2, a transcriptional target of Akt. NO also decreases expression of p63 that, like SOX2, is required for the development of stratified squamous epithelium. At the same time, NO increases the expression of CDX2, which can promote intestinal differentiation. Thus, NO triggers molecular events that inhibit squamous differentiation while promoting intestinal differentiation, a combination that might lead to development of the intestinal metaplasia of Barrett's oesophagus. These observations provide a rationale for research on interventions that might reduce NO production in the distal oesophagus as a way to prevent Barrett's oesophagus.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
- Data supplement 2 - Online table
Footnotes
KA and XH contributed equally.
Contributors All authors included in this paper fulfil the criteria of authorship and have approved the submission of this manuscript. Specific contributions are as follows: KA: study design; technical and material support; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript. XH: study design; technical and material support; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript. AA: study design; technical and material support; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript. XZ, CY, EC, QZ, KBD, THP, DHW, KI and TS: technical and material support; important intellectual content. RDO: analysis and interpretation of data; critical revision of manuscript; important intellectual content. SJS: study concept; analysis and interpretation of data; critical revision of manuscript; important intellectual content. RFS: study concept/design; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript.
Funding This work was supported by Merit Review Award #BX002666 from the US Department of Veterans Affairs Biomedical Laboratory Research Program (SJS), the National Institutes of Health (R01-DK63621 and R01-CA134571 to RFS and SJS.; K12 HD-068369-01 and K08-DK099383 to EC; R01-DK097340 to D.H.W.)
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.