Article Text

Abstract
Objective MicroRNAs (miRNAs) are well-known regulators of disease pathogenesis and have great potential as biomarkers and therapeutic targets. We aimed at profiling miRNAs in alcoholic hepatitis (AH) and identifying miRNAs potentially involved in liver injury.
Design MiRNA profiling was performed in liver samples from patients with AH, alcohol liver disease, non-alcoholic steatohepatitis, HCV disease and normal liver tissue. Expression of miRNAs was assessed in liver and serum from patients with AH and animal models. Mimic and decoy miR-182 were used in vitro and in vivo to evaluate miR-182's biological functions.
Results MiRNA expression profile in liver was highly altered in AH and distinctive from alcohol-induced cirrhotic livers. Moreover, we identified a set of 18 miRNAs predominantly expressed in AH as compared with other chronic liver conditions. Integrative miRNA-mRNA functional analysis revealed the association of AH-altered miRNAs with nuclear receptors, IGF-1 signalling and cholestasis. Interestingly, miR-182 was the most highly expressed miRNA in AH, which correlated with degree of ductular reaction, disease severity and short-term mortality. MiR-182 mimic induced an upregulation of inflammatory mediators in biliary cells. At experimental level, miR-182 was increased in biliary cells in mice fed with 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet but not upregulated by alcohol intake or fibrosis. Inhibition of miR-182 in DDC-fed mice reduced liver damage, bile acid accumulation and inflammatory response.
Conclusions AH is characterised by a deregulated miRNA profile, including miR-182, which is associated with disease severity and liver injury. These results highlight the potential of miRNAs as therapeutic targets and biomarkers in AH.
- ALCOHOLIC LIVER DISEASE
- BILIARY EPITHELIUM
- LIVER CIRRHOSIS
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Alcoholic hepatitis (AH) develops in patients with underlying alcoholic liver disease (ALD) and heavy alcohol intake and has a high short-term mortality. The pathogenesis of AH is still poorly understood; it is characterised by inflammatory cell infiltration, hepatocellular damage, steatosis, fibrosis, ductular reaction expansion and cholestasis.
MicroRNAs (miRNAs) are small non-coding RNA known to participate in the regulation of important pathophysiological pathways in liver diseases. miRNAs have been shown to be good diseases biomarkers as well as novel therapeutic targets.
MiR-182 is involved in organogenesis, cancer development and metastasis in several types of cancer.
What are the new findings?
MiRNA profiling in liver tissue revealed that AH is characterised by a specific miRNA expression profile differentiated from other chronic liver diseases including alcoholic-induced cirrhosis.
Integrative miRNA-mRNA functional analysis revealed the association of AH-altered miRNAs with fatty acid metabolism, bile acid transport and cholestasis.
MiR-182 is the most highly expressed miRNA in liver tissue in patients with AH and its expression correlates with disease severity, markers of ductular reaction and outcome.
MiR-182 is mainly expressed in ductular reaction cells and hepatocytes and mediates hepatocellular injury and promotes inflammation. Blockade of MiR-182 in experimental animal models reduces liver injury, bile acid accumulation and expression of pro-inflammatory genes.
How might it impact on clinical practice in the foreseeable future?
Understanding the molecular mechanisms regulating the pathogenesis of AH is of utmost importance for the development of new therapies in AH. In this study, we provide the first miRNA profile in AH and we identify individual miRNA potentially involved in AH pathogenesis. Importantly, we show that miR-182 is highly expressed in AH and correlates with disease severity and important features of AH. Moreover, experimental studies suggest its involvement in liver injury, inflammation and bile acid accumulation. All together, this study highlights the importance of miRNAs in AH and emphasises the potential of miRNAs as new therapeutic targets as well as future biomarkers.
Background and aims
Alcoholic liver disease (ALD) is a major cause of mortality worldwide and a major indication for liver transplantation in Europe and North America. Alcoholic hepatitis (AH) appears in patients with underlying ALD and heavy alcohol intake. Severe forms of AH still have a high short-term mortality, mainly due to incomplete response of patients to current available treatments (ie, corticosteroids and pentoxifylline).1 The pathogenesis of AH is still poorly understood; it is characterised by inflammatory cell infiltration, hepatocellular damage, steatosis, fibrosis, ductular reaction and cholestasis.2 Moreover, several histological features have been associated with disease severity and poor outcome in AH, including the degree of fibrosis, neutrophil infiltration, bilirubinostasis and ductular reaction.3 ,4
MicroRNAs (miRNAs) are small non-coding RNA of ∼ 22 nucleotides, which are post-transcriptional regulators of gene expression by targeting the 3′ untranslated region of the mRNA.5 MiRNA-target mRNA pairing leads to cleavage or translation inhibition depending on total or partial complementarity.6 ,7 The interaction of miRNA with target mRNA is complex; while each miRNA is able to recognise and target several gene transcripts, these can also be targeted by several miRNAs.8 ,9 Moreover, the gene and miRNA expression profile depends on the tissue, cell type or pathophysiological condition; therefore, the formation of miRNA-mRNA pair requires not only the complementarity but also the coexistence in time and place of both molecules. Liver diseases such as non-alcoholic steatohepatitis (NASH), hepatocellular carcinoma (HCC), HCV liver disease, ALD and biliary diseases are reportedly associated with important changes in miRNAs expression.10–15 These studies highlight the importance of miRNAs to understand the pathophysiology of liver diseases and to develop future therapies. However, there is very limited information regarding the expression of miRNAs in AH and their role in the pathogenesis of liver disease.
In the present study, we have identified a profile of miRNAs expressed in AH. By performing an integrative analysis of miRNA and mRNA data, we have shown that altered miRNAs in AH are associated with the regulation of key features of AH. Additionally, we selected miR-182 for further analysis based on its expression in AH, predicted targeted genes and potential biological functions. MiR-182 expression was high in AH, associated with disease severity and involved in liver injury.
Materials and methods
Patients
Samples were selected from a cohort of consecutive patients with clinical, analytical and histological features of AH admitted to the Liver Unit of the Hospital Clinic of Barcelona from July 2009 to January 2014. Forty-eight liver samples from patients with AH were selected based on total RNA availability and quality, and were used to perform miRNA array and hepatic miRNA expression validation. The clinical and biochemical parameters of patients with AH included in this study are shown in table 1, with no differences between the two cohorts (miRNA array cohort (n=13) and validation cohort (n=35)). Inclusion criteria for patients with AH were excessive alcohol consumption (>60 g/day) previous to admission, elevated levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), γ-glutamyltranspeptidase, bilirubin and histological diagnosis of AH. Liver samples of patients were obtained by transjugular biopsy at admission. On the other hand, exclusion criteria included the presence of HCC or other associated potential causes of liver disease other than alcohol consumption. Severe AH was defined as a Maddrey's discriminant function >32 and/or an age-bilirubin-international normalised ratio-creatinine (ABIC) score ≥6.71 at admission.16 Patients with severe AH were treated with prednisone orally 30 mg/day for 4 weeks followed by a 2-week taper period. During hospitalisation, patients were treated according to current international and local guidelines. All patients received nutritional as well as psychological assistance to help achieve alcohol abstinence.
Comparisons of demographic, clinical and haemodynamic characteristics between patients with alcoholic hepatitis in the microarray cohort and validation cohort
Patients with other aetiologies included in the study are further described in online supplementary material.
Supplemental material
Array studies
High-quality RNA samples from 13 AH, 5 ALD-induced cirrhosis (ALD-CH), 5 NASH-induced cirrhosis (NASH-CH), 4 HCV-induced cirrhosis (HCV-CH) and 6 normal livers were hybridised on a GeneChip miRNA 3.0 Array (Affymetrix, Santa Clara, California, USA) as described in online supplementary material. MiRNA array data have been deposited in NCBI's Gene Expression Omnibus with accession number GSE59492 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token=mbuxqyqmfdghrwd&acc=GSE59492).
MiRNA-mRNA profile integration and functional analysis
Among all samples analysed by miRNA array, eight AH and four normal liver samples were previously assessed for gene expression microarray analysis.17 Both paired miRNA and mRNA expression profiles were used for the integrative analysis by MiRComb package, as previously described.18 Functional analysis of significant dysregulated gene targets was performed with Ingenuity Pathway Analysis (Qiagen; Red Wood, California, USA). Canonical pathways and category disease annotation with p<0.05 were considered.
Determination of miR-182 in serum samples
Serum samples of 28 patients with AH and 8 control individuals were used to extract total RNA enriched in small RNAs with miRNeasy Serum/Plasma kit (Qiagen). Spike-in control (cel-miR-39a-3p) was added before extraction to later normalisation in the absence of an endogenous control. cDNA synthesis and qPCR were performed using miR-X miRNA First-Strand Synthesis Kit (Takara Bio Europe/Clontech, Saint-Germain-en-Laye, France) following manufacturer's instructions.
Mouse models
Animal models representing features of AH were performed: (1) ductular reaction model obtained with 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet; (2) ethanol intake; (3) fibrosis induced by carbon tetrachloride (CCl4) treatment and (4) acute-on-chronic liver injury mimicked by the induction of chronic damage with CCl4 plus acute ethanol (see online supplementary material). Animal procedures were conducted according to European Union regulations and were approved by the Ethics Committee of the University of Barcelona.
In vivo inhibition of miR-182 through a miRNA decoy in mice with DDC diet
Decoy systems to inhibit miRNA functions have been developed during the past 10 years.19 ,20 Among others, tough decoys (TuD) are plasmid constructs with hairpin-shape structures containing an internal loop with miRNA-binding sequences able to capture miRNA into stable complexes. Each decoy would be able to block a specific miRNA depending on the binding sequence.
Decoy to inhibit miR-182 and decoy control miR-1306 were developed by Dr. Brown Laboratory.21 Plasmid vectors of decoy-control or decoy-182 were cloned in Escherichia coli and were provided as bacterial stabs. E. coli was grown in Luria Broth medium overnight at 37°C. Thereafter, plasmid vectors were purified using PureLink Hipure Plasmid Maxiprep Kit (Life Technologies). To perform an in vivo delivery of decoys, plasmids were complexed with transfection reagent Jet PEI (PolyPlus, Illkirch, France) following the manufacturer's instructions. CD57/BL6 male 8-week-old mice were injected intraperitoneally with 50 µg of plasmid every 48 h during 1 week, with a total of four injections. Also, 24 h after the first injection mice were fed with 0.1% DDC diet or chow diet. Animals were sacrificed after 7 days of diet, blood and livers were collected.
In vitro miR-182 mimic transfection
H69 human cholangiocytes cell line (a gift from Dr. D. Jefferson, Tufts University, Boston, Massachusetts, USA), primary mouse hepatocytes and a macrophage mouse cell line (RAW 264.7) were transfected with 50 nM of hsa-miR-182 or mmu-miR-182, respectively (Exiqon, Vedbaek Denmark). Further information is detailed in online supplementary material.
Statistical analysis
Statistical analysis of the miRNA expression among groups and their correlation with clinical parameters and patient mortality were performed as described in online supplementary material.
Results
MiRNA expression profile in AH
In order to evaluate whether miRNA expression is altered in AH, we performed a wide miRNA profile analysis by microarray in 13 samples of liver tissue from patients with AH and 6 normal liver fragments. Unsupervised clustering analysis grouped AH and normal samples separately, showing the difference in miRNA expression between groups (p<0.05). The miRNA expression array revealed an important dysregulation of miRNA expression in AH compared with normal liver. In total, 111 miRNAs were upregulated in AH while 66 miRNAs were downregulated in AH versus normal samples (figure 1A). To validate the results from the microarray, we evaluated the hepatic expression of miRNAs known to be involved in liver diseases but not described in AH such as miR-21, miR-155, miR-214 and miR-422a by qPCR in a confirmatory cohort of patients with AH (n=35) and normal liver fragments (n=8). MiR-21, miR-155 and miR-214 were confirmed to be significantly upregulated and miR-422a downregulated in AH (figure 1B).

Profile of microRNA (miRNA) in alcoholic hepatitis (AH). (A) Heat map of the most differentially regulated miRNAs in patients with AH (n=13) compared with normal livers (n=6) (p<0.05). Red-coloured pixels correspond to an increased abundance of the miRNA in the indicated sample, whereas green pixels indicate decreased miRNA levels. (B) Validation by qPCR of four dysregulated miRNA in a confirmatory cohort of AH liver tissues (n=35) compared with normal liver (n=8). Results are expressed as mean±SD. (C) Multidimensional scaling analysis representing the 33 samples that underwent miRNA array profiling. The different samples are placed in the three-dimensional space according to their miRNA expression. AH samples are represented in dark blue (n=13), alcoholic liver disease-induced cirrhosis (ALD-CH) in green (n=5), HCV-CH in pink (n=4), non-alcoholic steatohepatitis (NASH)-CH in light blue (n=5) and normal livers in orange (n=6). (D) Heat map of the 18 miRNA specifically dysregulated in AH with respect to other liver diseases. In total, 17 were found to be increased in AH and only 1 was downregulated (p<0.05).
MiRNA expression profile in AH and other chronic liver disease
Next, we aimed at exploring whether altered miRNAs were exclusively expressed in AH (an acute-on-chronic condition) or are commonly altered in other chronic liver diseases. The miRNAs expression profile was assessed in liver samples from cirrhotic patients with ALD-CH and also NASH-CH and HCV-CH.
A multidimensional scaling analysis performed with all the samples analysed by miRNA array showed that samples belonging to the AH group were homogeneously distributed, with a wide separation from the other groups (figure 1C). These results suggest that AH express a profile of miRNAs different from chronic liver diseases, including ALD. As shown in figure 1D, 18 miRNAs were differentially expressed in AH compared with the other liver diseases analysed. Interestingly, among them, 17 miRNAs were upregulated and only 1 was downregulated. It is important to note that the set of upregulated miRNAs in AH showed a low level of expression in all the other aetiologies, suggesting that they may be regulating specific pathways underlying AH.
Importantly, a paired comparison of AH samples with samples from patients with ALD-CH showed that the miRNA expression profile clearly differentiated between the two groups, with 51 miRNAs differentially regulated in AH versus normal and ALD-CH (see online supplementary figure S1).
Altogether, these results suggest that AH is characterised by a specific miRNA profile, which differentiates AH from other chronic liver diseases.
To determine which genes may be regulated by the 18 differentially expressed miRNAs in AH, we performed an integrative analysis with Mircomb Package combining data from the miRNA profile and mRNA transcriptome from the same AH samples. The software selected miRNA-mRNA pairs that fulfilled the following two conditions; first, they presented a significant negative correlation between miRNA and gene expression, and second the miRNA-mRNA pair presented a predicted sequence complementarity. Remarkably, a functional analysis of genes potentially targeted by the 18 miRNAs expressed in AH, uncovered their relationship with pathways related to nuclear receptors PXR, RXR or FXR and also with hepatic cholestasis as shown in table 2.
Pathways regulated by the 18 microRNAs differentially expressed in alcoholic hepatitis
MiR-182 expression correlates with disease severity and key features of AH
We selected miR-182 for further evaluation since it was the most highly expressed miRNA in AH. The expression of miR-182 was assessed by qPCR in a confirmatory cohort of AH patients (n=35), showing an increase of 3.12-fold (p=0.001) in AH versus normal livers. Moreover, expression of miR-182 was higher in AH compared with liver samples from patients with HCV-CH and NASH (figure 2A). However, expression of miR-182 was not significantly different in AH and ALD-CH liver samples (3.12±1.96 and 2.65±1.52 fold vs normal livers, respectively), suggesting that it is not exclusively expressed in AH condition.
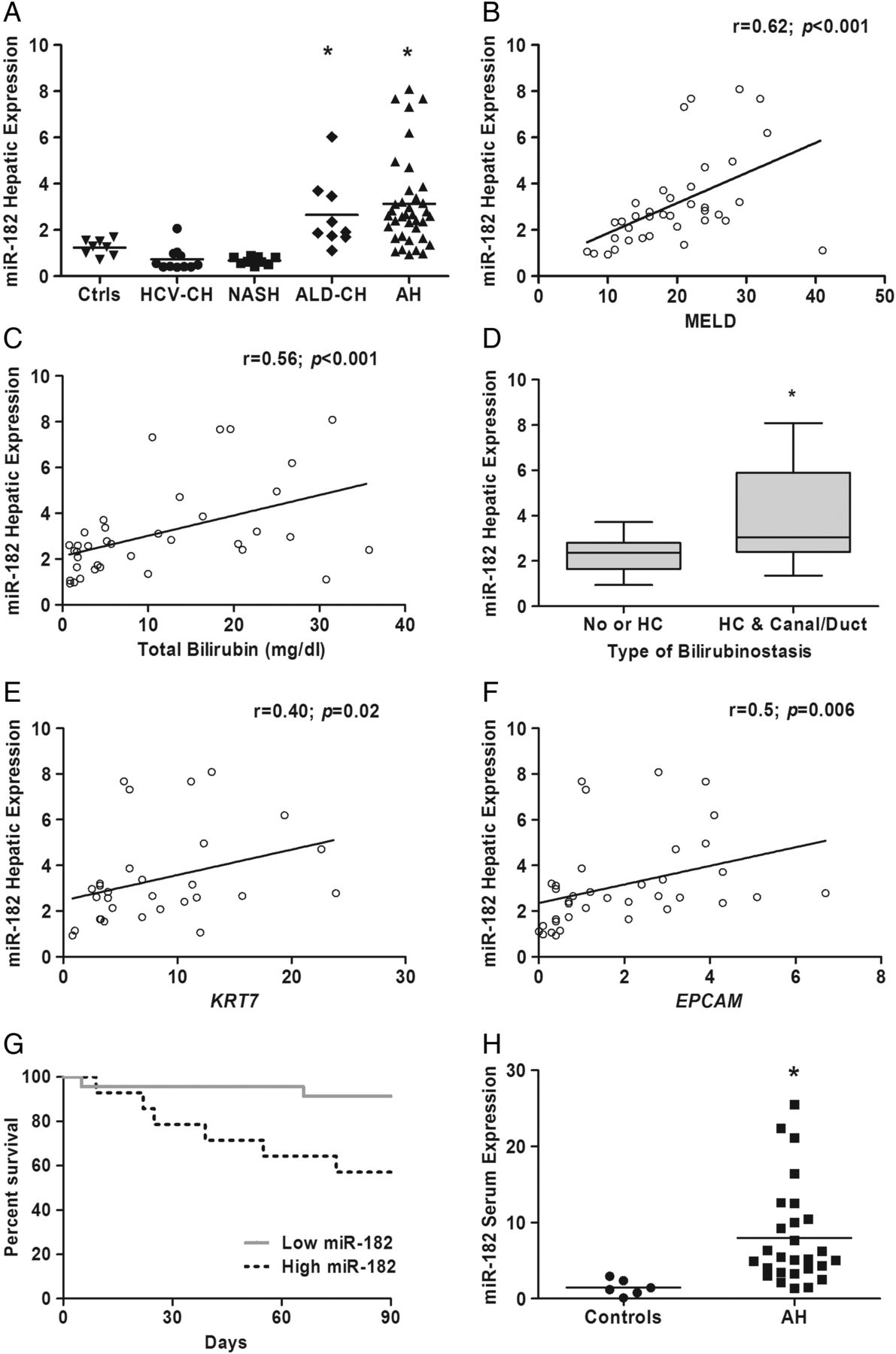
MiR-182 is increased in alcoholic hepatitis (AH) and correlates with scores of disease severity. (A) Hepatic expression of miR-182 assessed by qPCR in AH (n=35) compared with normal liver (n=8), HCV-cirrhosis (CH) liver (n=11), non-alcoholic steatohepatitis (NASH) liver (n=9) and ALD-induced CH (ALD-CH) liver (n=9) samples. *p<0.05 vs controls, HCV-CH, NASH. (B) Hepatic expression of miR-182 correlates with MELD score; and with (C) baseline total serum bilirubin (mg/dL). (D) Hepatic expression of miR-182 among different types of bilirubinostasis. HC, hepatocellular; Canal/Duct, canalicular and/or ductular. *p=0.01. Hepatic expression of miR-182 also correlates with hepatic expression of genes encoding liver progenitor cells (LPC) markers as (E) Keratin 7 (F) and EPCAM. (G) Kaplan–Meier analysis showing 90-day mortality according to miR-182 hepatic expression in patients with AH. A threshold of 2−ΔΔct=2.9 was determined as a cut-off value with the best sensitivity and specificity to define patients with low (2−ΔΔct<2.9) or high (2−ΔΔct≥2.9) hepatic expression of miR-182. (p=0.01). (H) miR-182 expression was determined in serum samples of patients with AH versus control individuals (p<0.05); expression was normalised with a Spike-In (cel-miR-39a-3p).
To further investigate the potential role of miR-182 in the pathogenesis of AH, we assessed whether its expression correlated with severity scores. We found a positive correlation between miR-182 hepatic expression and MELD score (p<0.001) (figure 2B) and also ABIC (p=0.005) (see online supplementary figure S2). Moreover, expression of miR-182 correlated with serum bilirubin levels (p<0.001) (figure 2C) and bilirubinostasis evaluated by histological assessment as previously described3 (p=0.01) (figure 2D). Interestingly, there was a positive correlation between miR-182 expression and keratin 7 and epithelial cell adhesion molecule (EPCAM), two markers of ductular reaction known to be associated with disease severity4 (figure 2E, F).
Next, we evaluated the association of miR-182 expression with short-term mortality of patients with AH. MiR-182 expression was higher in those patients who died within 90 days after admission (see online supplementary figure S2). Moreover, Kaplan–Meier analysis using a cut-off with the highest sensitivity and specificity (2.9-fold (2−ΔΔct)) compared with normal tissue (sensitivity 79%, specificity 74%; area under the receiver operating characteristic (AUROC) curve 0.76, 95% CI (0.56 to 0.96); p=0.01) showed the association of miR-182 expression above 2.9 with short-term mortality (figure 2G). Finally, to assess whether miR-182 could be detected circulating, we performed a qPCR in serum samples from patients with AH (n=28) versus control individuals (n=8). As shown in figure 2H, we found a marked increased in the expression of miR-182 in AH serum samples over normal individuals (p<0.05). Interestingly, miR-182 serum levels were not altered at 7 days after corticosteroid treatment in patients with AH. Moreover, miR-182 serum level did not correlate with liver tissue levels and was not associated with disease severity or mortality in patients with AH (see online supplementary figure S3), suggesting that at serum level it may not be a good biomarker in AH.
MiR-182 expression in animal models of liver injury
Although recent publications described animal models showing some of the features of severe ALD,22 ,23 no described animal model reproduces all the clinical features of AH, including an important degree of fibrosis, inflammation, ductular reaction and cholestasis. Therefore, miR-182 expression was evaluated not only in animal models of ethanol intake, but also models of fibrosis and ductular reaction alone and in combination with ethanol intake. As shown in figure 3A, B, no differences were found in hepatic expression of miR-182 in animals treated with CCl4 or ethanol alone. Conversely, a combined treatment of CCl4 plus ethanol induced a mild, although significant increase in miR-182 expression (1.68-fold, p<0.05) (figure 3B). Remarkably, in mice treated with DDC diet, which induces liver injury, inflammation, ductular reaction and cholestasis, miR-182 was upregulated in a time-dependent manner (figure 3C). However, combination of DDC with ethanol intake did not change the miR-182 expression (figure 3D). MiR-182 was not upregulated in an animal model of NASH (16 weeks feeding with high-fat diet) or in animals with acute T-cell-mediated liver injury (acute concanavalin A-treated animals, data not shown).

Differential hepatic expression of miR-182 in animal models of liver injury. MiR-182 hepatic expression in (A) mice treated with carbon tetrachloride (CCl4) (n=4) for 2 weeks; (B) mice treated with single ethanol gavage (n=3) or mice administered with CCl4 plus ethanol gavage (n=4); (C) mice treated with 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet during 1 week (n=5), 4 weeks (n=5) and 6 weeks (n=3) compared with chow-fed mice (n=3); (D) mice fed with Lieber-de-Carli diet supplemented with ethanol (n=5), DDC (n=3) or ethanol plus DDC (n=6). To investigate the contribution of cells types to miR-182 expression, (E) non-parenchymal cells (n=3) and ductular reaction cells (n=3) were compared with whole liver (n=3). Whole livers and cell fractions were obtained from mice fed with DDC diet for 3 weeks (*p<0.05 vs whole liver; #p<0.05 vs whole liver DDC; $p<0.05 vs negative fraction). (F) Hepatocytes were isolated from liver of mice treated with DDC diet during 6 weeks and miR-182 expression was assessed. Results expressed as mean±SD.
To assess the cell source of miR-182, parenchymal and non-parenchymal cells were isolated from mice fed with DDC diet. As shown in figure 3E, miR-182 was enriched in ductular reaction cells compared with whole liver tissue and not expressed in non-parenchymal cells (p<0.05). Although to a lesser effect, miR-182 was also expressed in hepatocytes (figure 3F), indicating that in injured liver miR-182 is expressed predominantly in ductular reaction cells and also in hepatocytes.
Functional analysis of miR-182
In order to evaluate the potential role of miR-182 in liver injury, we blocked miR-182 function in DDC-treated mice by using a miR-182 decoy (decoy-182). MiRNA decoys are constructs that act as sponges binding to mature miRNAs and preventing the pairing to target mRNA. Healthy mice treated with decoy-182 or decoy-control did not show changes in liver transaminases and biochemical parameters (figure 4A). Importantly, administration of decoy-182 in DDC-treated animals induced a reduction of liver injury as shown by a decrease of ALT, AST and lactate dehydrogenase (LDH) serum levels compared with decoy-control (figure 4A). Moreover, decoy-182 decreased serum alkaline phosphatase (AP) levels as well as total bile acids in liver tissue, suggesting that miR-182 may be involved in bile acids metabolism and cholestasis (figure 4A,B). As shown in figure 4C, expression levels of key inflammatory genes such as Mcp-1, Ccl20, Cxcl5, Cxcl1 or related to apoptosis like Bcl2 were reduced in DDC-treated animals that received decoy-182 compared with decoy-control. Decoy-182 treatment in DDC induced a non-significant increase in Slc1A1 expression and did not change the expression of Cfl-1 in whole liver tissue (figure 4C). Liver histology and the extension of ductular reaction were assessed by a blinded pathologist. However, no major changes were observed in neutrophil infiltration (figure 4D). Because miR-182 was mainly expressed in ductular reaction, we selected the H69 human cholangiocyte cell line to assess the role of miR-182 in vitro. Transfection of H69 with a miR-182 mimic induced a downregulation of predicted targets SLC1A1 and Cofilin1 and an upregulation of inflammatory and cell cycle genes such as CCL20, CXCL1, IL-8 and Cyclin D1 (figure 5A). Moreover, transfection of primary hepatocytes and macrophages with a miR-182 mimic induced a downregulation of the predicted target gene Cofilin1 and an upregulation of interleukin (IL)-6 in both cell types but did not change the expression of most inflammatory genes (figure 5B, C).

Inhibition of miR-182 in 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet mouse model. Mice were fed with chow or DDC diet during 1 week and both groups were treated with four injections of a decoy-control or a decoy-182. (A) Aspartate aminotransferase (AST), alanine aminotransferase (ALT), lactate dehydrogenase (LDH) and alkaline phosphatase (AP) serum levels were measured. (B) Levels of total bile acids measured in whole liver. (C) Hepatic gene expression measured by quantitative PCR and expressed as 2−ΔΔCt. (D) Representative images of haematoxylin/eosin staining and inmunohistochemistry of Keratin 19 and myeloperoxidase (MPO) in livers of DDC-fed mice treated with decoy-control or decoy-182. (E) Quantification of MPO-positive areas. Significances are shown above bars: *p<0.05; ns, non-significant.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mimicking the effects of miR-182 in vitro. Genes related to injury, inflammation and potential target genes of miR-182 were evaluated by qPCR in (A) H69 human cholangiocytes, (B) primary mouse hepatocytes and (C) a mouse macrophage cell line RAW 264.7. *p<0.05 vs cells transfected with scramble). Scr, scramble.
All together, these results suggest that miR-182 may be involved in cholangiocyte and liver injury and promote liver inflammation.
Discussion
In this study, we show that AH is characterised by an important change in miRNA expression. We compared the miRNA profile in different liver diseases to identify miRNAs that may be involved in the pathogenesis of AH. As a key differentially expressed miRNA, we identified miR-182, which expression correlates with disease severity and patient outcome. Experimental analysis of miR-182 in vitro and in vivo suggests that miR-182 may be involved in liver cell injury and inflammation. Moreover, integrative analysis of dysregulated miRNAs in AH and the role of their target genes suggest that key features of AH may be highly regulated by miRNAs, thus pointing out miRNAs as good targets to modulate the pathophysiology of AH.
In this study, we showed a global view of the miRNA landscape in AH. Moreover, we identified individual miRNAs with high probabilities of playing an important role in AH. In this regard, we highlighted several miRNAs differentially expressed in AH. While most miRNAs identified have never been described in the context of liver disease, others such as miR-21, miR-155 and miR-214 have already been reported as regulators of key pathways in liver diseases. MiR-21 is known to regulate liver fibrosis24–26 and regeneration.27 ,28 MiR-155 has been described to be expressed in inflammatory cells in alcohol-induced liver disease29 and in animal models of alcohol intake.30 Moreover, a recent study described miR-214 as a regulator of liver fibrogenesis by exosomal intercellular communication.31
An important finding of this study is the identification of a set of miRNAs dysregulated in AH compared with chronic liver diseases. The inclusion of patients with ALD-CH, HCV-CH and NASH-CH in this study has yielded important information regarding which genes particularly involved in AH pathogenesis may be regulated by miRNAs. Interestingly, the computational integrative analysis with miRNAs and gene expression data suggested that miRNAs in AH may be regulating genes involved in bile acid metabolism, transport and xenobiotic metabolism. By microarray analysis, we identified 51 miRNAs potentially deregulated in AH compared with ALD-CH and normal liver. However, the expression of miR-182 in patients with AH and ALD-CH evaluated by qPCR did not yield statistically significant differences. These results may be due to the fact that decompensated patients with ALD-CH have high levels of AP, bilirubinostasis and ductular reaction, parameters that correlate with miR-182 expression. Further studies should assess the expression of key miRNAs during the progression of ALD in bigger cohorts of patients in order to identify miRNAs specifically expressed in AH.
From all the miRNAs evaluated, we selected miR-182 for further evaluation based on its high expression in AH and its correlation with severity scores, including the outcome of patients with AH. Little is known about the role of miR-182 in liver physiology and disease.13 ,32 ,33 MiR-182 has been mainly described to be involved in cancer development and metastasis in several types of cancer,34–37 including HCC and liver metastasis.38 ,39 In the context of carcinogenesis, miR-182 has been reported to promote cell invasion, cell proliferation and angiogenesis. In liver, in vitro data suggest that miR-182 may have a role mainly in cholangiocytes, promoting the expression of inflammatory mediators, but also in hepatocytes and macrophages in which miR-182 induce the expression of IL-6. Notably, miR-182 has been proposed as a biomarker since several studies report its presence in blood circulation.40–42 Although miR-182 can be found increased in blood in patients with AH, the poor correlation with disease severity scores and patients outcome suggest that may not be useful as a biomarker in the context of AH.
The lack of an appropriate animal model that reproduces the main features of AH precluded the assessment of the role of miR-182 in a single animal model. To overcome this limitation and to identify the possible mechanisms behind the induction of miR-182 upregulation, we undertook two approaches. First, we evaluated the correlation of miR-182 expression with histological and clinical data from patients with AH; and second, we evaluated its expression in different animal models of liver injury, each of which is a well-established model of key features of liver injury and resolution such as fibrosis, ductular reaction and cholestasis, inflammation or alcohol toxicity. Both clinical data and the data obtained from animal models suggested that miR-182 is associated with cholangiocyte damage and ductular reaction, all relevant features in AH and known to be directly related to AH pathogenesis and patient outcome.3 ,4 Moreover, we also found miR-182 to be mainly expressed in ductular reaction cells.
In order to explore the effects of miR-182 in vivo, we performed a loss-of-function study using a decoy for miR-182. In recent years, DNA-encoded miRNA decoys have been used to understand the role of miRNAs in vitro and in vivo in different disease models.43–45 This approach is particularly interesting since it allows the long-term specific blockage of the desired miRNA. Treatment of DDC-fed animals with decoy-182 induced an important reduction of transaminases as well as LDH serum levels, suggesting the implication of miR-182 in liver injury. Interestingly, decoy-182 treatment had a profound effect on bile acid content and cholestasis since both, total bile acids in liver tissue and serum levels of AP, a surrogate marker of bile acid retention and a well-known marker of cholestasis, were reduced in animals treated with decoy-182. Moreover, bile acid accumulation can have a cytotoxic effect causing liver injury and inflammation.46–48 Therefore, reduced liver injury and inflammation observed in mice treated with decoy-182 could be an indirect effect resulting from the reduction of bile acid liver levels. Whether miR-182 is targeting genes involved in bile acid metabolism and transport or it is only indirectly associated with bile acid content will deserve further investigation. Although in vitro and in vivo studies with miR-182 suggest that interfering miR-182 may be a promising target in AH, further studies are required to better understand its role in liver disease. Moreover, we have to be cautious when proposing an miRNA as potential target for therapy since miRNAs are highly pleiotropic and expressed in several tissues and conditions.
This study does not provide information regarding the possible role of most of the miRNAs identified in AH. While some miRNAs might participate in the pathogenesis of the disease, others may be just markers associated with clinical features of AH. Interestingly, miRNAs have a great potential as biomarkers in the context of liver diseases, especially when detected in body fluids12 ,49 ,50; additional studies are necessary to evaluate whether other miRNAs besides miR-182 may be found in body fluids and can be used as non-invasive biomarkers of patient prognosis or for diagnostic purposes in AH.
Understanding the molecular mechanisms regulating the pathogenesis of AH is of utmost importance for the development of new therapies for AH. In 2012, we published the first transcriptome analysis of liver tissue from patients with AH, describing the main pathways dysregulated in this condition.17 We have now expanded our understanding of the regulation of AH by describing an miRNA profile in AH and identifying miRNA involved in AH pathogenesis. Global miRNA changes in alcoholic liver diseases, together with the identification of key individual miRNAs that may regulate AH, will certainly stimulate our understanding of this disease, the development of new therapeutic strategies and the identification of biomarkers.
Acknowledgments
This work was performed in the Centre Esther Koplowitz. The authors wish to thank Cristina Millán for her excellent technical support. For their advice and support, the authors wants to thank Dr Silvia Affò and Dr Oriol Morales-Ibanez. They are indebted to the Genomics Unit and Cytometry and Cell Sorting Facility of the Institut d'Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS) for their technical help. The human cholangyocites cell line (H69) was kindly provided by Dr Doug Jefferson, Department of Integrative Physiology and Pathobiology, Sackler School, Tufts University School of Medicine, Boston, MA. The keratin 19 (TROMA-III) antibody was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biology (Iowa City, IA). JA is part of the National System of Researches (SNI) and wishes to express his gratitude to the Mexican National Council of Science and Technology (CONACyT, Mexico City, Mexico) for partially supporting his predoctoral stay at IDIBAPS.
References
Footnotes
Contributors DB performed the experiments, collected and analysed the data, interpreted the data and wrote the manuscript. MV-C generated the integration package miRComb and performed the integration analysis. MC, DR-T, ML, LP and BA collected and interpreted data and contributed to critical revision of the manuscript. JA and IG collected human samples and analysed the data. JJL generated the integration package miRComb and interpreted the data of the microarray. AD performed histological analysis of human and mouse samples. JMB, JC, RB, JC and PG helped to design some parts of the study and interpreted the data and contributed to critical revision of the manuscript. PS-B conceived and designed the study, interpreted the data and wrote the manuscript.
Funding This work was supported by grants from Fondo de Investigación Sanitaria Carlos III (FIS), co-financed by Fondo Europeo de Desarrollo Regional (FEDER), Unión Europea, ‘Una manera de hacer Europa’ (FIS PI14/00320, PI12/00330, FIS PI12/01265 and FIS PI11/01990 to PS-B, PG, JC and RB, respectively) and from the NIH (1U01AA021908-01-33490 to PS-B and 1U01AA021908 to RB). PS-B is funded by Instituto de Salud Carlos III, Miguel Servet (CP11/00071 and CON14/00129) and co-financed by Fondo Europeo de Desarrollo Europeo (FEDER), Unión Europea, ‘Una manera de hacer Europa’. PG is funded by Agencia de Gestió d'Ajuts Universitaris i de Recerca (AGAUR) 2014 SGR 708, Centro de Investigación Biomédica en Red de Enfermedades Hepaticas y Digestivas (CIBEReHD) and Institució Catalana de Recerca i Estudis Avançats (ICREA). MC and IG are funded by Instituto de Salud Carlos III, by Sara Borrell and Rio Hortega grants respectively. MV-C and DR-T received a grant from the Ministerio de Educación, Cultura y Deporte, FPU programme.
Competing interests None declared.
Patient consent Obtained.
Ethics approval Ethics Committee of the Hospital Clinic of Barcelona.
Provenance and peer review Not commissioned; externally peer reviewed.