Article Text

Abstract
Objective Dietary fibre has beneficial effects on energy metabolism, and the majority of studies have focused on short-chain fatty acids produced by gut microbiota. Ginseng has been reported to aid in body weight management, however, its mechanism of action is not yet clear. In this study, we focused on the potential modulating effect of ginseng on gut microbiota, aiming to identify specific strains and their metabolites, especially long-chain fatty acids (LCFA), which mediate the anti-obesity effects of ginseng.
Design Db/db mice were gavaged with ginseng extract (GE) and the effects of GE on gut microbiota were evaluated using 16S rDNA-based high throughput sequencing. To confirm the candidate fatty acids, untargeted metabolomics analyses of the serum and medium samples were performed.
Results We demonstrated that GE can induce Enterococcus faecalis, which can produce an unsaturated LCFA, myristoleic acid (MA). Our results indicate that E. faecalis and its metabolite MA can reduce adiposity by brown adipose tissue (BAT) activation and beige fat formation. In addition, the gene of E. faecalis encoding Acyl-CoA thioesterases (ACOTs) exhibited the biosynthetic potential to synthesise MA, as knockdown (KD) of the ACOT gene by CRISPR-dCas9 significantly reduced MA production. Furthermore, exogenous treatment with KD E. faecalis could not reproduce the beneficial effects of wild type E. faecalis, which work by augmenting the circulating MA levels.
Conclusions Our results demonstrated that the gut microbiota-LCFA-BAT axis plays an important role in host metabolism, which may provide a strategic advantage for the next generation of anti-obesity drug development.
- obesity
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Ginseng extract (GE) has an anti-obesity effect.
The activation of thermogenesis in brown adipose tissue (BAT) and beige fat is beneficial for weight management.
Modulation of gut microbiota using prebiotics and probiotics may improve host metabolism and reduce obesity.
What are the new findings?
GE exhibited an anti-obesity effect by activating BAT.
GE specifically increased the abundance of E. faecalis in the gut and E. faecalis administration reduced body weight through BAT activation.
E. faecalis facilitated its beneficial effects on metabolism through its metabolites; the most important one being myristoleic acid (MA), a long-chain fatty acid.
Acyl-CoA thioesterase is at least partially responsible for the biosynthesis of MA in E. faecalis.
How might it impact on clinical practice in the foreseeable future?
E. faecalis, as a novel anti-obesity probiotic, can be exploited to manage excess weight and obesity.
MA is a new postbiotic from E. faecalis with beneficial effects on obesity and its related diseases.
Introduction
Obesity has become a global epidemic. It leads to an increased risk of various diseases including insulin resistance, type 2 diabetes, fatty liver disease, cardiovascular disease and certain types of cancers.1 2 Obesity develops due to higher energy intake than energy expenditure, which results in excess energy storage in white adipose tissue (WAT) in the form of triglycerides.3 The current anti-obesity strategies are mainly aimed at restricting energy uptake and absorption. However, obesity is far from being dealt with satisfactorily. Previous studies have shown that brown adipose tissue (BAT) facilitates weight control, health, and provides an anti-obesity effect.4 Therefore, increasing BAT activity could be a novel and effective therapeutic approach for preventing and curing obesity and its related diseases.5–7
Intestinal flora is an important environmental factor for energy acquisition and storage in obesity, and thus, it has an impact on the host.8 9 Interestingly, there are reports demonstrating that acclimation to cold enhances host metabolism by modulating gut microbiota.10 Therefore, strategies that modulate the gut microbiota have been proposed to prevent and treat obesity. Previous studies have shown that modulation of gut microbiota using prebiotics and probiotics may improve host metabolism and reduce obesity and associated metabolic diseases. Examples include the effect of polysaccharides and dietary fibres in reducing body weight and alleviating type 2 diabetes by inducing specific probiotics and their derived metabolites including short-chain fatty acid (SCFA) and probiotic strains such as Lactobacillus and Bifidobacterium, which can attenuate obesity comorbidities.11 12 In addition, it has been reported that long-chain fatty acids (LCFAs), which are defined as saturated and/or unsaturated fatty acids with 14–20 carbons, also play important roles in regulating the energy metabolism.13 There are reports that eicosapentaenoic acid induces thermogenesis in brown adipocytes through FFAR4-dependent upregulation of miR-30b and miR-378,14 and myristic acid improves hyperglycaemia in a mouse model of type 2 diabetes.15 Besides, LCFAs, which are produced in brown adipocytes by the lipolysis, can activate uncoupling protein 1 (UCP1) in thermogenic fat.15 Importantly, a recent study indicated that LCFAs are essential for UCP1 uncoupling.16
Traditional Chinese Medicine (TCM), often administered orally, has been widely used in the treatment of various diseases and dates back to thousands of years. TCM (such as Ganoderma lucidum) can modulate the gut microbiota composition by increasing probiotics and reducing pathogens, thus preventing the development and progression of obesity and other diseases.17 However, its mechanism of action is not yet clear. Panax ginseng is widely used as a traditional medicine or functional food.18 19 The ginsenosides are the major active components of Panax ginseng, responsible for various biological functions including effects against obesity and obesity-related diseases,20 and are metabolised to rare saponins through deglycosylation reactions by the gut microbiota before being absorbed into the blood (eg, ginsenoside CK).21 22 In addition, a study has shown that long-term administration of ginseng extracts (GEs) could change the gut microbiota composition.23 Therefore, further studies are needed to better understand the complex interactions between ginseng and gut microbiota and secondary metabolites and obesity.
In the present study, we revealed that GE significantly increased energy expenditure and reduced adiposity via BAT activation. Furthermore, GE induced the intestinal bacterium E. faecalis, which can produce myristoleic acid (MA), which reduces obesity through increasing BAT activity and beige fat formation. Additionally, we found that the Acyl-CoA thioesterase (ACOT) gene of E. faecalis has a potential for MA biosynthesis. Thus, our data indicated that E. faecalis and its metabolite, MA, might be used for the prevention and treatment of obesity and its complications.
Methods
Mice
Four-week-old male C57BLKS/J-Leprdb/Leprdb (db/db) mice were purchased from the Model Animal Research Center of Nanjing University. Four-week-old male C57BL/6J mice were obtained from Vital River Laboratory Animal Technology (Beijing, China). UCP1 luciferase transgenic mice (ThermoMouse) were purchased from Jackson Labs (Bar Harbor, Maine, USA) and backcrossed with C57BL/6J mice. The UCP1 KO mice (genetic background C57Bl/6J) were originally established by L Kozak (Pennington Medical Research Centre, Baton Rouge, Louisiana, USA). WT and UCP1 KO mice were obtained from heterozygous breeding pairs in our SPF laboratory animal-house (Institute of Zoology, Chinese Academy of Sciences, Beijing, China) and the genotype was evaluated as described previously.24 Each group of mice was fed a chow diet or a high-fat diet (HFD; 60% kcal fat as indicated, Beijing HuaFuKang Bioscience, Beijing, China). GE (10 mg/kg), bacterial strains (1010 CFU/day, Aoke Biology Research, Beijing, China), or fatty acids (5 mg/kg, ANPEL Laboratory Technologies, Shanghai, China) were administered by oral gavage during the indicated period starting from the age of 4–6 weeks. For in vivo antibiotic treatment, db/db mice were treated with combined antibiotics (ABX) (containing 100 µg/mL neomycin (Sigma), 50 µg/mL streptomycin (Sigma), 100 U/mL penicillin (Sigma), 50 µg/mL vancomycin (Sigma), 100 µg/mL metronidazole (Sigma), 1 mg/mL bacitracin (Sigma), 125 ug/mL ciprofloxacin (Sigma), 170 ug/mL gentamycin (Sigma) and 10 ug/mL chloramphenicol (Lablead, Beijing, China)) in sterile water for 5 days. After ABX treatment, the ACOT WT and KD E. faecalis strains were fed by oral gavage for 1 week (1010 CFU/day).
All the mice were housed in groups of 4–5 animals per cage in a pathogen-free facility with a 12 hours: 12 hours light: dark cycle with ad libitum access to food and water. In all the experiments, the mice were fasted overnight before euthanasia using mild ether anaesthesia, and sacrificed. All the animal studies were approbated by the Institutional Animal Care and Use Committee of Institute of Zoology (Chinese Academy of Sciences).
Analysis of gut microbiota
The gut microbiota was analysed as previously described.22 Briefly, genomic DNA (0.25 g) was isolated from bacterial colonies with the PowerSoil DNA Isolation Kit (MO BIO Laboratories, USA). After amplification and purification of the V3-V4 region of bacterial 16 s rRNA genes, the abundance and diversity of intestinal flora in mice were determined using Illumina HiSeq sequencing (Novogene, Beijing, China). The raw reads have been submitted to the BIGD Genome Sequence Archive database (Accession number CRA001467). Sequencing libraries were assessed utilising Agilent 2100 bioanalyzer (Agilent Technologies, USA) and the qualified libraries were amplified on cBot to obtain the gene clusters on a flow-cell.
According to the unique barcodes, paired-end reads were allocated to the samples. The overlapped regions between paired-end reads were combined using FLASH v1.2.7. According to the QIIME (V1.7.0) quality control process, high-quality clean tags were obtained by qualitatively filtering the raw reads under specific filtering conditions. Based on the UCHIME algorithm, the chimaera sequences were detected by comparing tags with the Gold database and then removed. The QIIME software package was used to conduct the bioinformatic analyses of the sequences. Sequences sharing at least 97% comparability were attributed to the same operational taxonomic units (OTUs). QIME was used to carry out alpha, beta diversity and Principal Coordinates Analysis depending on the unweighted unifrac distances.
Establishment of ACOT gene knockdown strain of E. faecalis
The ACOT gene of E. faecalis was knocked down using the CRISPR Interference System according to the method established by Qi.25 The dCas9 gene was cloned from the plasmid pHR-SFFV-dCas9-BFP-KRAB (46911, Addgene). The sgRNA scaffold with J23119 promotor was synthesised (Genscript, Nanjing, China). The genes were amplified and cloned into the vector SlpA −8148 (a gift from Dr Yanling Hao) containing a constitutive promoter SlpA and a chloramphenicol-selectable marker. The ACOT gene knockdown (KD) strain was created by electro-transformation with dCas9-expressing and ACOT-targeting sgRNA (sequence: ATCCCTTTGGTGCACTATT) vector, while the WT strain was created by electro-transformation with a dCas9-expressing and sgRNA scaffold vector. Chloramphenicol (10 µg/mL) was used to select the positive clones. After culturing in MRS medium at 37°C overnight, the total RNA was isolated using a bacteria RNA Extraction Kit (R403-1, Vazyme, Nanjing, China) and cDNA was synthesised with HiScript III first Strand cDNA Synthesis Kit (R312-01, Vazyme, Nanjing, China). The ACOT gene expression level was detected as described above, while 16S rDNA was chosen as an internal reference.
Statistics
Data are expressed as means±SE. Statistics were performed using a repeated measure two-way analysis of variance (ANOVA), the one-way ANOVA test, analysis of covariance (ANCOVA) or Student’s t-test. Statistical significance was set at p<0.05. *p<0.05; **p<0.01; ***p<0.0001.
Results
GE decreases adiposity and enhances energy metabolism by BAT activation
In this study, we assessed the possibility of the anti-obesity effects of GE mediated by gut microbiota and BAT. GE (10 mg/kg) was administered to db/db mice by daily oral gavage. After 8 weeks, their body weight gain were significantly decreased compared with those receiving the vehicle treatment (figure 1A). The body composition was analysed using CT. Consistent with body weight change, the percentage of whole-body fat decreased by 11% compared with that of the control mice (figure 1B). Moreover, the organ weight of epididymal fat was significantly decreased after GE treatment (figure 1C), whereas the organ weight of BAT, subcutaneous fat (Sub), and liver did not show significant changes (figure 1C). These results indicated that GE treatment significantly reduced body weight and adiposity in db/db mice. Adiposity is often accompanied with an alteration of the energy balance. Consequently, we examined whether GE treatment could affect energy metabolism via indirect calorimetry (using a TSE labmaster system). Interestingly, GE treatment significantly increased energy expenditure compared with the control mice (figure 1D), while there was no significant difference in the food intake or energy intake (online supplementary figure 1A,D), physical activity (online supplementary figure 1B), or respiratory exchange ratio (RER) (online supplementary figure 1C), absorbed energy (online supplementary figure 1E) and no absorbed energy (online supplementary figure 1F) between the two groups. The results of energy balance strongly indicated that the anti-obesity role of GE is due to increased energy expenditure rather than reduced food intake or increased physical activity. Furthermore, GE treatment greatly increased core body temperature when animals were exposed to a cold environment (4°C, 4 hours) (figure 1E,F). Importantly, GE treatment improved glucose homoeostasis (online supplementary figure 1G) and insulin sensitivity(online supplementary figure 1H), and reduced hepatic steatosis (online supplementary figure 1I–K). These results indicated that GE treatment improved the energy homoeostasis in obese mice. This can be partially explained by the improved BAT thermogenic activity (figure 1G) and the increased expression of UCP1, which generates heat through the uncoupling process and OXPHOS expression in BAT and Sub of the GE treated db/db mice (figure 1H,I). BAT is a thermogenic organ, essential in maintaining core body temperature and its thermogenic activity is inhibited at thermoneutrality.26 However, GE treatment did not affect body weight gain at thermoneutrality (online supplementary figure 1L). This result proved that the anti-obesity role of GE certainly depends on the BAT activity. Taken together, these results indicated that GE increased whole body energy metabolism by BAT activation without altering the energy intake or physical activity.
Supplemental material

Ginseng extract (GE) increases whole-body energy metabolism and BAT activity in db/db mice. Db/db mice were treated with GE (10 mg/kg) for 8 weeks by daily oral gavage (n=7–8). (A–C) GE treatment significantly decreased body weight gain as well as the EP organ weight of db/db mice. (D–F) Energy expenditure and core body temperature were significantly increased after GE treatment in db/db mice. (G–I) GE treatment significantly increased UCP1 expression and mitochondrial oxphos protein expression in BAT and Sub of db/db mice. For body weight curves, statistical analysis was performed using a repeated measure two-way ANOVA, for energy expenditure, ANCOVA were employed and the rest of the statistics was performed with Student’s t-test. Data are means±SEM *p<0.05; **p<0.01; NS, not statistically significant. ATP, adenosine triphosphate; BAT, brown adipose tissue; EP, epididymal fat; SDHB, succinate dehydrogenase complex,subunit B; UCP, uncoupling protein; UQCRC, ubiquinol cytochrome c reductase core protein.
GE treatment enriches E. faecalis
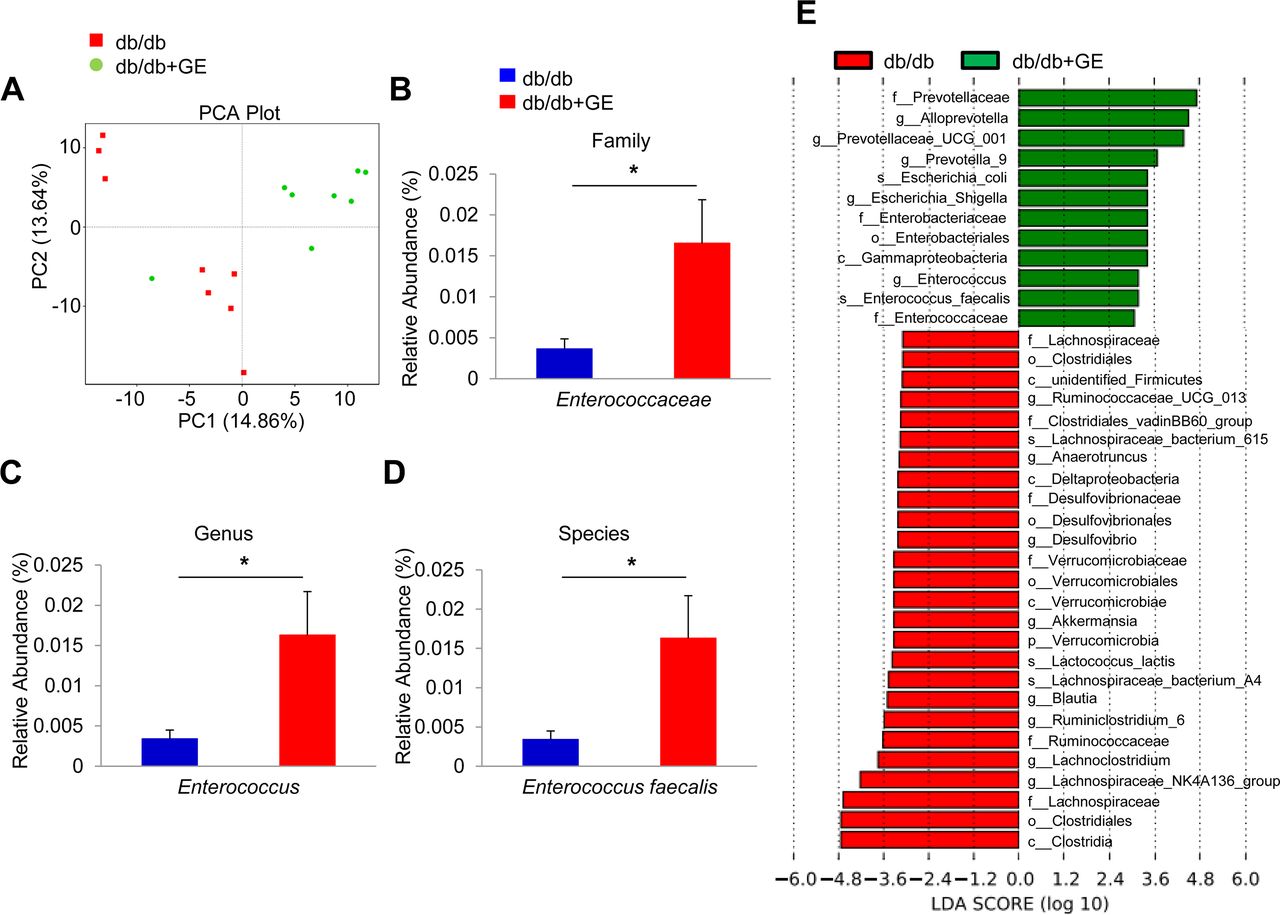
It was previously demonstrated that ginsenosides can be deglycosylated by the actions of the gut microbiota.27 Therefore, we speculated on whether gut microbiota could play key anti-obesity roles in GE-treated mice using this mode of action. Towards this end, faecal samples were examined for bacterial community diversity via 16S rRNA gene amplicon sequencing. The α-diversity values for the bacterial community analysis with ACE, Chao 1, Shannon index and Simpson index in the GE treated mice were significantly lower than those in the control mice (online supplementary table 1). The overall structure of the gut microbiota that was investigated by principal component analysis showed that the two groups were clearly differentiated (figure 2A). The phylum-level proportional abundances, especially within Firmicutes and Bacteroidetes, did not show significant differences after GE treatment (online supplementary figure 2A). This might be due to the limitations of the chosen technology; the 16S rRNA gene amplicon sequencing could not cover 100% of the gut microbiota. Beyond this, there were 35 OTUs that were significantly altered by GE in db/db mice (online supplementary figure 2B). Interestingly, we found that E. faecalis was considerably enriched at the family-genus-species levels (figure 2B–D). Thus, our results indicated that the composition of gut microbiota changed significantly in response to GE treatment and that E. faecalis was substantially enriched (figure 2E).
Supplemental material
Supplemental material

Ginseng extract (GE) alters microbiota composition in db/db mice. Db/db mice were treated with GE (10 mg/kg) for 8 weeks by daily oral gavage (n=8). (A) Plots shown were generated using the weighted version of the UniFrac-based principal component analysis. (B–D) Comparison proportion of family, genus and species levels of E. faecalis in faeces detected by pyrosequencing analysis. (E) Discriminative taxa determined by LEfSe between two groups (log10 LDA >3.5). Statistical analysis was performed using Student’s t-test between db/db and db/db+GE groups. Data are means±SEM *p<0.05. LDA, linear discriminant analysis,
E. faecalis administration reduces obesity
E. faecalis is a gram-positive bacterium belonging to Lactobacillales at the order level. E. faecalis is a common commensal complement to the healthy human gastrointestinal tract as well as a leading cause of hospital-acquired infections.28 To investigate the role of E. faecalis in obesity, we used HFD-fed mice as our model. Interestingly, we found a significant reduction both in body weight gain and fat mass in the mice treated with E. faecalis (figure 3A,B). We next examined whether E. faecalis treatment could affect energy metabolism. Notably, E. faecalis treatment significantly increased the core body temperature when animals were exposed to a cold environment (4°C, 4 hours) (figure 3C,D). In addition, in the E. faecalis-treated mice, energy expenditure increased significantly compared with that of the control mice (figure 3E). Consistently, histological analysis revealed that the sizes of lipid droplets in BAT from the E. faecalis-treated mice were much smaller than those of the control mice (figure 3F). Correspondingly, the protein expressions of UCP1 and OXPHOS were significantly upregulated in BAT and Sub adipose tissue of the E. faecalis-treated mice (figure 3G,H). Furthermore, E. faecalis treatment improved glucose homoeostasis and reduced hepatic steatosis (online supplementary figure 3D–H). These beneficial effects were not significantly related to the difference in food intake, physical activity, or RER (online supplementary figure 3A–C). Importantly, E. faecalis treatment did not affect body weight gain at thermoneutrality (online supplementary figure 3I). Thus, these results strongly indicated that E. faecalis increased energy expenditure and reduced adiposity by activating BAT and beige fat formation.
Supplemental material

E. faecalis (EF) increases whole-body energy metabolism and BAT activity in HFD-fed mice. HFD-fed mice were treated daily with EF (1×1010 CFU) by oral gavage for 8 weeks (n=7–9). (A–B) EF treatment significantly decreased body weight gain as well as the fat mass of HFD-fed mice. (C–E) Core body temperature and energy expenditure were significantly increased after EF treatment in HFD-fed mice. (F–H) EF treatment significantly increased UCP1 expression and mitochondrial oxphos protein expression in bat and sub of HFD-fed mice. For body weight curves, statistical analysis was performed using a repeated measure two-way ANOVA, for energy expenditure, ANCOVA were employed and the rest of the statistics was performed with Student’s t-test. Data are means±SEM.*p<0.05; **p<0.01; ***p<0.0001; NS, not statistically significant. ATP, adenosine triphosphate; BAT, brown adipose tissue; EP, epididymal fat; HFD, high-fat diet; HFD+EF, high-fat diet with E. faecalis; SDHB, succinate dehydrogenase complex,subunit B; UCP, uncoupling protein; UQCRC, ubiquinol cytochrome c reductase core protein.
MA is enriched in serum metabolites and its administration reduces adiposity
A potential mechanism by which E. faecalis influences the host energy metabolism would be via the secondary metabolites generated during microbial fermentation in the gut. It was reported that UCP1 is activated by LCFAs.29 Recently, it has been reported that LCFAs serve as substrates permanently attached to UCP1 that promote the transfer of H+ from the inner to the outer mitochondrial membrane.16 Hence, it was of interest to know if E. faecalis treatment alters serum LCFAs. Accordingly, we performed an untargeted metabolomics analysis of LCFAs in the serum samples. Oral gavage of E. faecalis significantly altered a variety of LCFAs (figure 4A). To identify which of these LCFAs were specifically altered after E. faecalis treatment, we further analysed the amount of LCFAs in the culture medium with and without E. faecalis treatment. Intriguingly, we found a very high amount of MA (C14:1) in the E. faecalis treatment that reached a maximum of 12.3 times the amount found without the treatment (figure 4B). In addition, non-adecylic acid (C19:0) and linoleic acid (C18:2) also showed some degree of significant accumulation (5.9 and 4.3 times the level without the treatment, respectively), whereas this accumulation pattern was found to a lesser extent with other LCFAs (figure 4B). These results strongly suggested that E. faecalis may be involved in fatty acid metabolism. To further investigate the potential effect of LCFAs on BAT activity, primary brown adipocytes were treated with various LCFAs and cellular oxygen consumption was assessed. The results of the cellular experiment demonstrated that the top-ranking metabolite, MA, significantly up-regulated oxygen consumption (figure 4C). However, other fatty acids did not show such effects, including linoleic acid (online supplementary figure 4A). Moreover, MA significantly increased UCP1 expression (online supplementary figure 4B). These results implied that MA might increase the BAT activity. To further clarify this hypothesis, we used transgenic mice in which luciferase activity reveals endogenous UCP1 expression.30 Interestingly, the luciferase activity was significantly increased on MA treatment (online supplementary figure 4C,D). These results evidenced the potential of the efficacy of MA against obesity. To test this hypothesis, db/db mice were fed MA by oral gavage. As expected, MA significantly reduced body weight gain (figure 4D) and adiposity (figure 4E, online supplementary figure 4E) via increasing BAT thermogenesis and whole-body energy metabolism (figure 4F,G, online supplementary figure 4F). In addition, we revealed that MA activated BAT (figure 4H,I), induced beige fat formation (figure 4J), and reversed hepatic steatosis (online supplementary figure 4J–K). These beneficial effects did not significantly correlate with alterations in either food intake, or physical activity, or RER (online supplementary figure 4G–I). To assess whether our results might be explained by the anti-obesity effect of MA on BAT, we used UCP1 KO mice, which display greatly reduced BAT activity, to further verify our results. Accordingly, MA was able to reduce the gain in body weight induced by an HFD in wild type (WT) mice, but not in UCP1 KO mice (online supplementary figure 5A–C). In addition, MA could increase BAT thermogenesis and improve glucose homoeostasis under HFD conditions in WT mice (due to hypothermia, the cold challenge experiment was not performed on UCP1 KO mice) (online supplementary figure 5D–G). Additionally, energy expenditure increased in the WT mice, but not in UCP1 KO mice (online supplementary figure 5H). Furthermore, MA improved BAT hypertrophy and reversed hepatic steatosis in WT mice, but not in the UCP1 KO mice (online supplementary figure 5K, L, O, P). Accordingly, the UCP1 protein expression was significantly up-regulated in BAT and Sub of the MA treated WT mice (online supplementary figure 5M–N). Furthermore, these beneficial effects were not mediated by food intake or physical activity (online supplementary figure 5I–J). Importantly, MA treatment did not affect body weight gain at thermoneutrality (online supplementary figure 5Q). Taken together, these results demonstrated that E. faecalis-produced MA-reduced adiposity by increasing BAT activity and whole-body metabolism.
Supplemental material
Supplemental material

Myristoleic acid (MA) administration reduces adiposity. (A) E. faecalis (EF) treatment alters serum LCFAs in HFD-fed mice (n=9). (B) EF increases MA levels in MRS broth (n=6). (C) Oxygen consumption rates at day 6 of brown adipogenesis with 10 uM MA or BSA treatment (n=5–6). To investigate the potential effects of MA on whole-body energy metabolism, db/db mice were treated with MA (5 mg/kg) for 8 weeks by daily oral gavage (n=7–10). (D–E) MA treatment significantly decreased body weight gain as well as the fat mass of db/db mice. (F–G) Core body temperature and energy expenditure were significantly increased after MA treatment in db/db mice. (H) Representative H&E staining of BAT from db/db mice. (I–J) MA treatment significantly increased UCP1 expression and mitochondrial OXPHOS protein expression in BAT and Sub of db/db mice. For body weight and seahorse oxygen consumption curves, statistical analysis was performed using a repeated measure two-way ANOVA, for energy expenditure, ANCOVA were employed and the rest of the statistics were performed with Student’s t-test. Data are means±SEM. *p<0.05, **p<0.01. ATP, adenosine triphosphate; BAT, brown adipose tissue; EP, epididymal fat; HFD, high-fat diet; LCFA, long-chain fatty acid; SDHB, succinate dehydrogenase complex,subunit B; UCP, uncoupling protein; UQCRC, ubiquinol cytochrome c reductase core protein.
Reduction of E. faecalis MA production impairs anti-obesity effects
The previous report demonstrated that ACOTs are a large family of enzymes that catalyse the hydrolysis of the thioester bond between a carbonyl group and a sulfur atom, producing fatty acids and CoA (figure 5A). In humans, Them1 thioesterase hydrolyzes a range of fatty acyl-CoAs with a preference for long-chain acyl-CoA molecules, and dimerisation is induced by fatty acyl-CoAs, coenzyme A (CoASH).31 To elucidate how E. faecalis is involved in MA production, we first successfully established ACOT gene knockdown (KD) and WT E. faecalis strain using the Crispr-dCas9 technique (about 60% KD compared with WT) (figure 5B). KD of ACOT gene did not alter the growth of E. faecalis (data not shown). Next, the WT and KD E. faecalis strains were incubated in a De Man, Rogosa and Sharpe (MRS) culture medium, and LCFAs were subjected to an untargeted metabolomics analysis. Consistently, the WT strain was able to induce MA accumulation, whereas the ACOT KD strain could not induce as much accumulation of MA as the WT strain (figure 5C). In addition, KD of ACOT gene did not significantly alter other LCFAs or SCFAs compared with the WT E. faecalis (online supplementary figure 6A). These results clearly indicated that the production of MA is at least partly mediated by ACOT. To investigate the influence of genetically modified E. faecalis strains on UCP1 activity, a variety of mouse models were used. We observed that WT E. faecalis but not KD E. faecalis could reduce adiposity in HFD-fed mice (figure 5D,E, online supplementary figure 6B). Accordingly, WT but not KD E. faecalis was able to increase cold-induced thermogenesis (figure 5G, online supplementary figure 6C) and energy expenditure (figure 5F) without significant changes in food intake (online supplementary figure 6D). Endogenous UCP1 activity was also significantly upregulated in WT strain but not with the KD E. faecalis treatment in the UCP1 luciferase transgenic mice model (figure 5H,I). To avoid interspecies crosstalk between gut microbiota, mice were pretreated with ABX. Consistently, we found that the WT strain could increase BAT activity and cold-induced thermogenesis, whereas, the KD strain-treated db/db mice could not (online supplementary figure 6E–F). These results clearly demonstrated the important role of the ACOT gene in MA biosynthesis. Taken together, we showed that E. faecalis drove MA production via the ACOT gene.
Supplemental material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acyl-CoA thioesterase (ACOT) of the E. faecalis (EF) is a key enzyme involved in MA synthesis. (A) Synthesis pathway of free fatty acid by ACOT. (B) Successful knock down of ACOT gene in EF strain by Crispr-dCas9 technique (n=4–5). (C) Changes of LCFAs biosynthesis by WT and ACOT KD EF in MRS broth (n=4). HFD-fed mice were treated daily with WT and ACOT KD EF (1×1010 CFU) by oral gavage for 5 weeks (n=6–8). (D–E) Body weight gain and fat mass were significantly decreased after being treated with the WT strain but not with the KD strain in HFD-fed mice. (F–G) Energy expenditure and core body temperature were significantly increased after WT strain but not KD strain treated in HFD-fed mice. (H–I) Endogenous UCP1 activity was significantly decreased after being treated with the WT strain but not the kD strain in HFD-fed mice (n=5). (J) Ginseng extract enriched EF which can produce LCFA, specifically MA by ACOT gene and this further reduced obesity by increasing bat activity and beige fat formation. Data are means±SEM. HFD, high-fat diet. For body weight curves, statistical analysis was performed using a repeated measure two-way ANOVA, for energy expenditure, ANCOVA were employed and the rest of the statistics was performed by one-way ANOVA with Tukey’s post-hoc tests. *p<0.05; **p<0.01; ***p<0.0001; NS, not statistically significant. BAT, brown adipose tissue; EP, epididymal fat; HFD, high-fat diet; HFD+WT EF, high-fat diet with WT E. faecalis; HFD+KD EF, high-fat diet with ACOT KD EF; KD, knock down; LCFA, long-chain fatty acid; UCP, uncoupling protein; WT, wild type.
Discussion
Although it has been shown that as a TCM, ginseng displays various biological functions including effects against obesity and obesity-related diseases, the underlying molecular mechanism is still unclear. In this study, by integrating data on the gut microbiome, serum metabolome, and BAT, we demonstrated that the GE–E. faecalis−LCFA (specifically MA) axis reduces obesity by increasing BAT activity and beige fat formation. Moreover, E. faecalis was observed to drive the MA production via the ACOT gene. In addition, we demonstrated for the first time in this study that as an LCFA, E. faecalis-produced MA has tremendous potential for treating metabolic syndrome disorders, such as obesity (figure 5J).
Obesity is a chronic medical condition, which develops due to many factors, including genetic factors, lifestyle, diet, gut microbiota, and unbalanced energy metabolism. Therefore, drugs that aim to increase energy expenditure have the potential to be the most effective and attractive strategy against obesity.32 As an energy-consuming organ, BAT has received much attention.33 Therefore, increasing BAT mass and/or activity is a promising strategy to treat obesity and metabolic diseases. Indeed, studies by our group and others have shown that BAT transplantation or the use of rutin, which is able to activate BAT, could reverse metabolic disorders in various obese mouse models.34–37
Recently, several reports have pointed to the interaction between BAT and intestinal flora.10 These results suggest that GE-mediated brown fat activation might be associated with changes in the intestinal flora. Additionally, GE could also change the gut microbiota composition.23 Accordingly, we found that E. faecalis were significantly enriched at the family-genus-species levels after GE treatment. Furthermore, we found that E. faecalis could significantly reduce obesity by increasing BAT activity and beige fat formation. Interestingly, previous reports demonstrated that heat-treated E. faecalis (bacteria that were heated at 110°C for 10 min) had anti-obesity effects and improved hepatic steatosis in DIO mice model.38 A study reports that autoclaving Akkermansia muciniphila eliminated the anti-obesity effects of the live strain.39 In line with these observations, presumably, E. faecalis were biologically inactive after heat treatment, even though there is a lack of knowledge of the molecular mechanisms involved.
To understand the mechanisms of action, we speculated that the secondary metabolites that were produced during fermentation could play important roles in BAT activation. Interestingly, there are reports demonstrating that rats treated with SCFA, such as propionate and butyrate, showed significantly less gain in body weight than that seen in untreated rats, despite similar food intake.40 However, another study indicates that acetate promotes metabolic syndromes via the microbiome-brain-β-cell axis.41 Surprisingly, in our experiment, SCFAs showed minimal or no differences after E. faecalis treatment (online supplementary figure 6A). Interestingly, we found that LCFA (MA) production was exclusively elevated after E. faecalis treatment. Furthermore, fatty acids 19:0 and 18:2 also showed some degrees of increase after E. faecalis treatment (figure 4B). Recently, it has been reported that LCFAs serve as permanently attached UCP1 substrates that help to carry H+ via UCP1.16 The results of the oxygen consumption experiment indicated that C19:0 and C18:2 have minor effects compared with those of C14:1 (online supplementary figure 4A). These results highlighted the fact that E. faecalis induces MA levels, thereby involving UCP1-mediated heat generation. Detailed molecular analysis revealed that the E. faecalis-encoding ACOT gene was responsible for the biosynthesis of MA. In addition, in the treatment of the ACOT gene KD, E. faecalis was unable to reduce the body weight gain compared with that in the WT E. faecalis in HFD-fed mice. However, data on the types of substrates (eg, SCFAs or carbohydrates) that favour MA production are still largely unknown. Besides, our research does not rule out other possible alternative mechanisms of the anti-obesity effects of ginseng, including reduced adipocyte hypertrophy via the modulation of angiogenesis and MMP activity,42 increased energy expenditure via stimulation of the adenosine monophosphate-activated kinase pathway,20 reduced adipogenesis and inflammation,43 and maintenance of the gut barrier integrity.44
In summary, by integrating data on the gut microbiome, serum metabolome, and BAT, we demonstrated that the GE–E. faecalis–LCFA (specifically MA) axis reduces obesity by increasing BAT activity and beige fat formation. In addition, we observed that E. faecalis drives MA production via the ACOT gene. Furthermore, we demonstrated for the first time that as an LCFA, E. faecalis-produced MA has tremendous potential to treat metabolic syndrome disorders, such as obesity.
Acknowledgments
The authors would like to thank Dr Dangsheng Li for his great advices.
References
Footnotes
L-HQ, CZ, MD, JJ, HX and CY contributed equally.
Correction notice This article has been corrected since it published Online First. A second co-corresponding author has been added.
Contributors WJ, L-HQ, CZ, MD and DL. designed the research and analysed all of the results; L-HQ, JJ, HX and CY performed animal studies; CZ performed cellular experiments and animal tissue analysis; MD performed animal tissue analysis and bacteria dCas9 construction; XL, HZhou, HZhang, LC, Z-BL and F-LZ performed animal tissue analysis and whole-body animal imaging; S-ML and GS performed lipidomics analysis; and L-HQ, CZ, MD and WJ wrote the paper.
Funding This work was supported by the strategic priority research program (XDB13030000 to WJ), National Natural Science Foundation of China (31171131 and 81370951 to WJ, 81660643 to LQ), National Key Research and Development Program of China (2017YFC1001003 to WJ).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval All animal studies were approved by the In- stitutional Animal Care and Use Committee of Institute of Zoology (Chinese Academy of Sciences).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available in a public, open access repository. Data are available upon reasonable request.