Article Text

Abstract
Background and aims Inflammatory bowel disease (IBD) has been linked to a loss of tolerance towards the resident microflora. Therapeutic use of probiotics is known to be strain specific, but precise mechanisms remain unclear. The role of NOD2 signalling and the protective effect of Lactobacillus peptidoglycan (PGN) and derived muropeptides in experimental colitis were evaluated.
Methods The anti-inflammatory capacity of lactobacilli and derived bacterial compounds was evaluated using the 2,4,6-trinitrobenzene sulfonic acid (TNBS) colitis model. The role of NOD2, MyD88 and interleukin 10 (IL-10) in this protection was studied using Nod2−/−, MyD88−/− and Il10-deficient mice, while induction of regulatory dendritic cells (DCs) was monitored through the expansion of CD103+ DCs in mesenteric lymph nodes or after adoptive transfer of bone marrow-derived DCs. The development of regulatory T cells was investigated by following the expansion of CD4+FoxP3+ cells. High-performance liquid chromatography and mass spectrometry were used to analyse the PGN structural differences.
Results The protective capacity of strain Lactobacillus salivarius Ls33 was correlated with a local IL-10 production and was abolished in Nod2-deficient mice. PGN purified from Ls33 rescued mice from colitis in an IL-10-dependent manner and favoured the development of CD103+ DCs and CD4+Foxp3+ regulatory T cells. In vitro Ls33 PGN induced IL-10-producing DCs able to achieve in vivo protection after adoptive transfer in a NOD2-dependent way. This protection was also correlated with an upregulation of the indoleamine 2,3-dioxygenase immunosuppressive pathway. The protective capacity was not obtained with PGN purified from a non-anti-inflammatory strain. Structural analysis of PGNs highlighted in Ls33 the presence of an additional muropeptide, M-tri-Lys. The synthesised ligand protected mice from colitis in a NOD2-dependent but MyD88-independent manner.
Conclusions The results indicated that PGN and derived muropeptides are active compounds in probiotic functionality and might represent a useful therapeutic strategy in IBD.
- Probiotics
- peptidoglycan
- IBD
- NOD2
- dendritic cells
- inflammatory bowel disease
- mucosal immunity
- 2,4,6-trinitrobenzene sulfonic acid
Statistics from Altmetric.com
- Probiotics
- peptidoglycan
- IBD
- NOD2
- dendritic cells
- inflammatory bowel disease
- mucosal immunity
- 2,4,6-trinitrobenzene sulfonic acid
Significance of this study
What is already known about this subject?
The intestinal microbiota plays a major role in the development and maintenance of intestinal health.
Probiotic microorganisms might positively influence disease outcome of patients with inflammatory bowel disease (IBD). Effects remain strain specific and mechanisms of action are just starting to be unravelled.
NOD2 senses peptidoglycan-derived muropeptides, and mutations in Nod2 are associated with susceptibility to Crohn's disease (CD).
CD103+ dendritic cells (DCs) have been shown to drive preferentially CD4+Foxp3+ T regulatory cells notably through the indoleamine 2,3-dioxygenase (IDO)-dependent pathway, and as such play a crucial role in intestinal homesostasis.
What are the new findings?
The protective capacity of selected probiotic lactobacilli is NOD2-dependent and correlated with local interleukin 10 (IL-10) production.
Peptidoglycan (PGN) of lactobacilli can rescue mice from colitis in a strain-specific and IL-10-dependent way through induction of regulatory CD103+ DCs and regulatory T cells.
IDO expression is upregulated in the colon and in CD11c+ DCs purified from mesenteric lymph nodes in PGN-treated-mice.
The protective capacity of PGN of lactobacilli is linked to the presence of a NOD2 ligand exhibiting NOD2-dependent anti-inflammatory capacities.
How might they impact on clinical practice in the foreseeable future?
The use of probiotics in IBD is often based on empiric criteria, and several well-known probiotic strains failed to fulfil the expected clinical outcome, in particular in patients suffering from CD. Negative results can probably be linked to the fact that the probiotic selection relied mainly on gut survival or adhesion capacity rather than on immunological criteria such as their anti-inflammatory potential.
Our work clearly explained the strain specificity observed in the anti-inflammatory potential of selected lactobacilli, and indicated that the limited efficacy of lactobacilli in CD could be related to NOD2 deficiency. Although emerging evidence suggests that regulatory responses are crucial in circumventing inflammatory processes, our observations support an important role for probiotic and probiotic-derived components in regulating immune homeostasis. Insights gained through these findings will thus improve strain selection procedures and hopefully provide highly positive results in future clinical trials with probiotics in IBD. The identification of a functional molecule opens the route towards the design of a new family of therapeutic tools in such chronic inflammatory diseases.
Introduction
The microbiota is in permanent interaction with the host epithelium and immune system and contributes to gastrointestinal health.1 In genetically susceptible individuals, an inappropriate mucosal immune response against the intestinal flora appears to be the principal mechanism leading to the pathogenesis of inflammatory bowel disease (IBD). Such inappropriate immune responses are normally restrained by regulatory lymphocytes,2 among which CD4+Foxp3+ T cells play an important functional role, promoting tolerance to the flora and dietary proteins. A specialised population of intestinal dendritic cells (DCs) that express the integrin αE chain CD103 has recently been shown to promote the differentiation of naïve CD4+ T cells into Foxp3+ T regulatory (Treg) cells via a transforming growth factor β (TGF-β) and retinoic acid (RA)-dependent mechanism.3 Activation of nuclear factor-κB (NF-κB), a major regulator of inflammatory responses, is a key molecular event involved in the pathogenesis of IBD.4 Commensals can regulate such inflammatory responses by dampening this signalling pathway.5 6 However, a physiological level of inflammation, in response to activation of Toll-like receptors (TLRs) by commensals is also needed to maintain gut homeostasis.7 8 Previous studies indicated an altered microbial composition in the gut microbiota of patients with IBD.9 Probiotics, mainly represented by commensal lactic acid bacteria (LAB), are non-pathogenic living microorganisms which, when administered in sufficient amounts, confer beneficial health effects to the host (WHO/Food and Agriculture Organization (FAO) definition).10 Therefore, the use of probiotics to modulate the luminal contents in order to prevent or reduce intestinal inflammation represents an attractive and safe approach for treating IBD. Following positive results obtained with different animal models of colitis, clinical intervention studies indeed showed beneficial activities of some probiotics in patients suffering from pouchitis or ulcerative colitis.11–13 Their protective effects clearly depended on the strains used, and several well-characterised LAB strains failed to fulfil the expected clinical outcome, in particular in patients suffering from Crohn's disease (CD).14 We have previously shown that protection by LAB against colitis induced by TNBS (trinitrobenzene sulfonic acid) in mice was strain specific and correlated well with the in vitro immunomodulatory properties of the strains.15 More recently, we observed that some lactobacilli (eg, Lactobacillus salivarius Ls33) partially activated murine bone marrow-derived DCs (BMDCs), inducing regulatory DCs able to confer protection towards TNBS-induced colitis after adoptive transfer, in a NOD2-dependent manner.16 NOD2 is a member of the NOD-like receptor family, which recognises the smallest bioactive component of peptidoglycan (PGN) found in the bacterial cell wall, the muramyl dipeptide (MDP).17 Since mutations in the Card15 gene (coding for NOD2) are known to be involved in patients suffering from CD,18 we therefore evaluated if the beneficial effect of lactobacilli is mediated through PGN and the NOD2 signalling pathway.
We first confirmed that the protective properties of lactobacilli required NOD2 signalling. Consequently we demonstrated both in vitro and in vivo that PGN purified from the protective L. salivarius Ls33 strain exhibited anti-inflammatory potential, while PGN derived from a non-protective strain did not. We also showed that Ls33 PGN stimulated both DCs and T cell regulatory functions and linked its anti-inflammatory capacities to the presence of a specific NOD2 ligand.
Materials and methods
Bacterial strains and products
The two Lactobacillus strains L. salivarius Ls33 and L. acidophilus NCFM were obtained from Danisco (Madison,WI,USA). Bacteria were grown overnight at 37°C in de Man, Rogosa and Sharpe broth (Difco, Becton Dickinson, Sparks, MD, USA), washed twice in sterile phosphate-buffered saline (PBS) and resuspended in carbonate buffer (0.2 M NaHCO3, 2% glucose) or in PBS for in vivo and in vitro experiments, respectively. Lipopolysaccharide (LPS; Escherichia coli serotype 0111:B4) and Bacillus subtilis lipoteichoic acid (LTA) were purchased from Sigma (St. Louis, Mo, USA), and purified Staphylococcus aureus LTA from InvivoGen (San Diego, CA, USA). Ls33 LTA was prepared as previously described.19 20 PGNs were purified from the strains Ls33 and NCFM and their purity was evaluated as detailed in the Supplementary Materials and methods (see Supplementary figure 1). The muropeptide composition analysis was performed as described by Courtin21 (see Supplementary Materials and methods). Structural analysis of purified muropeptides by MALDI-TOF (matrix-assisted laser desorption/ionisation-time of flight) mass spectrometry was performed as previously described by Antignac et al.22 The M-tri-Lys M-tri-Lys (M-tri) and M-tri-Lys-d-Asn (M-tri-N) identified in PGNs of lactobacilli were chemically synthesised, purified by high-performance liquid chromatography (HPLC) and verified by H-nuclear magnetic resonance (NMR) and MALDI-TOF mass spectrometry at Cayla-InvivoGen. Their NOD2-sensing capacity was studied using a NOD2-expressing HEK293 cell line (see Supplementary Materials and methods).
Mice
BALB/c, wild-type (WT) and Il10-deficient C57BL/6J mice (7–9 weeks old) were purchased from Charles River (Oncins, France). MyD88 C57BL/6J-deficient mice (6–7 weeks old) were obtained from S. Akira23 through a generous gift of J.C. Sirard (INSERM U1019, Lille, France). Card15/Nod2-deficient (Nod2−/−) C57BL/6J mice (7–11 weeks old) were initially provided by J.-P. Hugot (Hôpital Robert Debré, Paris), further backcrossed in C57BL/6J to reach the eighth backcross by J. Viala (Institut Pasteur, Paris), as previously described.24 25
BMDCs generation and stimulation
DCs were generated from the bone marrow (BMDCs) of WT or Nod2−/− mice as previously described.16 For flow cytometry and ELISA analysis, BMDCs were incubated overnight with medium alone, LPS (100 ng/ml) or with PGN purified from Ls33 or NCFM (10 μg/ml). For in vivo adoptive transfer, BMDCs were stimulated overnight with PGN from Ls33 or NCFM (10 μg/ml), live Ls33 bacteria (multiplicity of infection (MOI) 10:1) in the presence of 150 μg/ml gentamicin or were left untreated. Cells were harvested by centrifugation, washed three times with sterile PBS and resuspended in PBS.
Experimental TNBS-induced colitis and study design
A standardised murine TNBS colitis model was used to induce acute levels of inflammation in mice.26 Briefly, anaesthetised BALB/c, WT and Il10-deficient C57BL/6J mice received an intrarectal administration of a 50 μl solution of the hapten reagent TNBS (Sigma-Aldrich, Saint Quentin Fallavier, France) dissolved in 50% ethanol (100 and 150 mg/kg for BALB/c and C57BL/6 mice, respectively). As Nod2−/− mice are more susceptible to colitis than their WT counterparts,24 two procedures were performed. Procedure 1: in order to obtain a similar level of inflammation in both mice lineages, Nod2−/− mice received a 40% lower dose of TNBS than WT mice (dose ranking from 120 to 170 mg/kg for Nod2−/− and WT mice, respectively). Procedure 2: both mice lineages received a dose of 145 mg/kg. The latter procedure was also used for Myd88−/− mice.
The anti-inflammatory properties of L. salivarius Ls33 were evaluated by the intragastric administration of live bacteria (2×108 colony-forming units (CFU)) or carbonate buffer to mice once daily for five consecutive days before colitis induction. The protective effect of PGN (50, 100 or 200 μg in 100 μl of PBS) was evaluated either by a single intraperitoneal administration concomitantly with the TNBS instillation or by oral feeding for 5 days before colitis induction. The protective effect of M-tri (10, 20 or 40 μg per mice), M-tri-N (20 μg per mice) and Ls33 LTA (100 μg per mice) was assessed by intraperitoneal administration (in 100 μl of PBS) just before TNBS instillation. The effect of Ls33- or PGN-treated BMDCs was studied by a single intraperitoneal administration of 2×106 cells just before TNBS administration. Animals were sacrificed 48 or 72 h after TNBS administration (for BALB/c and C57BL/6 mice, respectively). The evaluation of inflammation parameters was performed as described in detail in the Supplementary Materials and methods.
Total splenocytes and cells from mesenteric lymph nodes (MLNs) were stimulated at 2×106 cells/ml for 48 h with coated anti-CD3 (2 μg/ml; BD Biosciences, San Jose, CA, USA) and soluble anti-CD28 antibody (1 μg/ml; BD Biosciences). CD11c+ DCs were enriched from MLNs using CD11c (N418) microbeads (Miltenyi Biotec, Auburn, CA, USA). CD4+, CD4+CD25+ and CD4+CD25− T cells were isolated from MLNs with the CD4+ T cell isolation kit and the CD4+CD25+ regulatory T cell isolation kit (Miltenyi Biotec), respectively.
ELISA analysis
The concentrations of murine tumour necrosis factor α (TNFα), IL-10, IL-1β, CCL3 (chemokine ligand 3)/MIP-1α (macrophage inflammatory protein 1-α), CXCL2 (chemokine (C-X-C motif) ligand 2)/MIP-2, IL-12p70 and human TNFα were measured in culture supernatants or in colon extracts using specific R&D Duoset (R&D, Minneapolis, MN, USA) kits, according to the manufacturer's instructions.
Flow cytometry
Monoclonal antibodies (mAbs) against mouse CD4, FoxP3, CD11c, major histocompatibilty complex (MHC) II, CD80, CD40, CD86, CD103 and isotypes were purchased from eBioscience (San Diego, CA, USA) or BD Biosciences. For all staining, Fc receptors were blocked with the mAb anti-CD32 (2.4G2) as previously described.16 Flow cytometry was performed using a FACSCalibur and the CellQuest software (BD Biosciences).
Statistical analysis
Statistical analysis was performed using the non-parametric Mann–Whitney U test. Differences were considered to be statistically significant when the p value was <0.05.
Results
The anti-inflammatory properties of L. salivarius Ls33 require NOD2 signalling
To investigate the role of NOD2 signalling in the anti-inflammatory properties of L. salivarius Ls33, we assessed its protective effect in the TNBS colitis model using C57BL/6 WT and Nod2−/− mice. As Nod2−/− mice are more susceptible to colitis than WT mice,24 we first normalised the TNBS dose according to the weight of the mice in order to obtain similar levels of inflammation in both mice lineages (procedure 1). Three days following colitis induction, loss of body weight and strong macroscopic inflammation of the colon (Wallace score) were observed in both mice lineages (figure 1A). In WT mice, daily intragastric administration of Ls33 significantly reduced (p<0.001) these parameters (figure 1A), as well as the colonic levels of proinflammatory mediators (IL-1β and CXCL2), while leading to an increase of local IL-10 production (figure 1B). In contrast, the protective effect of the bacteria and the IL-10 production were abrogated in NOD2-deficient mice. When the same dose of TNBS was used in both mice lineages (procedure 2), inflammation was increased in Nod2−/− mice as compared with their WT counterparts. As before, while Ls33 was able to rescue WT mice from colitis, this was not observed in their Nod2−/−counterparts (Supplementary figure 2). Taken together, these data indicate that NOD2 signalling is involved in the anti-inflammatory properties of Ls33.

The protective effect of L. salivarius Ls33 against trinitrobenzene sulfonic acid (TNBS)-induced colitis is abrogated in Nod2-deficient mice. Wild-type (WT) or Nod2−/− C57BL/6 mice were fed for 5 days with either carbonate buffer (buffer) or 2×108 live L. salivarius Ls33 (Ls33) prior to TNBS administration (procedure 1). (A) Loss of body weight, Wallace score and (B) levels of colonic interleukin 1β (IL-1β), chemokine (C-X-C motif) ligand 2 (CXCL2) and IL-10 assessed 3 days after induction of colitis. Data represent the mean of 8–10 mice per group with the SEM (**p<0.01; ***p<0.001 vs buffer-treated WT mice).
Ls33 PGN protects mice from TNBS-induced colitis
As NOD2 detects degradation products of PGN,17 highly purified PGN was prepared from the anti-inflammatory strain L. salivarius Ls33. Its protective effect was assessed in the TNBS colitis model in BALB/c mice. Inflammation parameters (body weight loss, Wallace and Ameho scores and colonic myeloperoxidase (MPO) activity) were significantly reduced upon a single intraperitoneal injection of 100 μg of Ls33 PGN, conferring up to 60% protection according to the Wallace score (figure 2A). Histological examination of the distal colon of mice treated with TNBS showed destruction and necrosis of epithelial cells, thickening of the submucosa and an important infiltration of mononuclear cells, mainly represented by neutrophils. These inflammation parameters were significantly reduced upon Ls33 PGN administration (figure 2B). While TNBS instillation enhanced the transcription of the proinflammatory genes Il1b, Il6 and Tnf, injection of Ls33 PGN significantly reduced the transcription of these genes and, furthermore, enhanced the transcription of two genes involved in the immunosuppressive indoleamine 2,3-dioxygenase (IDO) pathway, Indo and Ifng, the latter encoding interferon γ (IFNγ) (figure 2C). While Ls33 PGN-treated mice showed a decrease of colonic Il10 gene expression (figure 2C), the protection was associated with an increased level of the IL-10 protein, measured in the colon and in the supernatant of stimulated splenic and MLN cells (figure 2D). The role of IL-10 was confirmed using Il10-deficient mice in which the protective effect of Ls33 PGN was abrogated (figure 2E). Finally, flow cytometric analysis revealed a 3.5-fold increase of regulatory CD11c+CD103+ DCs (figure 2F) and a significant enhancement of CD4+FoxP3+ Treg cells in the MLNs of Ls33 PGN-treated mice (figure 2H). This was matched by an increased Foxp3 mRNA expression in both CD4+CD25+ and CD4+CD25− T cells (figure 2I) as well as increased Ifng and indo gene expression in CD11c+ DCs (figure 2G). However, Ifng gene expression was not modulated in CD4+ purified T cells from PGN-treated mice (data not shown), indicating that this was not linked to a shift in the T helper 1 (Th1) response. Importantly, the protective effect of PGN could be achieved against inflammation of varying degrees of severity (Supplementary figure 3) and no protective effect was observed after intraperitoneal administration of Ls33 LTA (Supplementary figure 4).

Intraperitoneal administration of Ls33 peptidoglycan (PGN) protects mice against experimental colitis and induces CD103+ dendritic cells (DCs) and CD4+ FoxP3+ regulatory T cells in mesenteric lymph nodes (MLNs). (A–D) BALB/c mice were injected intraperitonally with phosphate-buffered saline (PBS) or 100 μg of Ls33 PGN concomitantly with trinitrobenzene sulfonic acid (TNBS) instillation. All parameters were assessed 2 days after colitis induction. Data represent the mean of 10 mice per group with the SEM (**p<0.01; ***p<0.001 vs PBS-treated mice). (A) Loss of body weight, macroscopic and microscopic colonic inflammation (Wallace and Ameho scores, respectively) and myeloperoxidase (MPO) activity. (B) Histological analysis of colonic sections after May–Grünwald Giemsa staining (original magnification, ×20). (C) Gene expression of proinflammatory and immunoregulatory molecules in colonic samples harvested 2 days after induction of colitis assessed by real-time PCR. (D) Levels of interleukin 10 (IL-10) in colon extracts or in culture supernatant of anti-CD3/CD28-stimulated cells isolated from spleen or MLNs 2 days after colitis induction. (E) Wild-type (WT) or Il10-deficient (Il10−/−) C57BL/6 mice were injected intraperitoneally with PBS or 100 μg of Ls33 PGN concomitantly with TNBS instillation. The Wallace score was assessed 2 days later. Data represent mean values of 7–9 animals. Error bars represent the SEM (**p<0.01 vs PBS-treated WT mice). (F–I) BALB/c mice were injected intraperitoneally with PBS or 100 μg of Ls33 PGN concomitantly with TNBS instillation, and phenotypic analysis of MLN cells was assessed 2 days after colitis induction. (F) CD11c+ DCs were enriched from murine MLNs and the expression of CD103 was evaluated by flow cytometry. Data are representative of two independent experiments (n=9–10 mice per group; **p<0.01 vs PBS-treated mice). (G) Levels of Ifng and indo mRNA in MLN-enriched CD11c+ DCs 2 days after TNBS administration measured by real-time PCR. Values are expressed as the relative mRNA levels compared with PBS-treated mice. Data represent mean values of 8 mice with the SEM (*p<0.05 vs PBS-treated mice). (H) Total CD4+ cells from murine MLNs were stained for CD4 and FoxP3. Representative percentages of double positive cells are shown in the dot plot. Data from six mice are gathered in the graph (*p<0.05 vs PBS-treated mice). (I) Relative expression of Foxp3 in MLN CD4+CD25+ or CD4+CD25− isolated T cells evaluated by real-time PCR. Data represent the mean values of three mice with the SD (*p<0.05 vs PBS-treated mice).
The anti-inflammatory properties of Ls33 PGN were also confirmed after oral administration (figure 3). Oral feeding with Ls33 PGN reduced, in a dose-dependent manner, the body weight loss, the macroscopic inflammation and the transcription of proinflammatory genes such as Cxcl2. Taken as a whole, these data highlight that PGN purified from an anti-inflammatory Lactobacillus strain is able to protect mice from colitis.
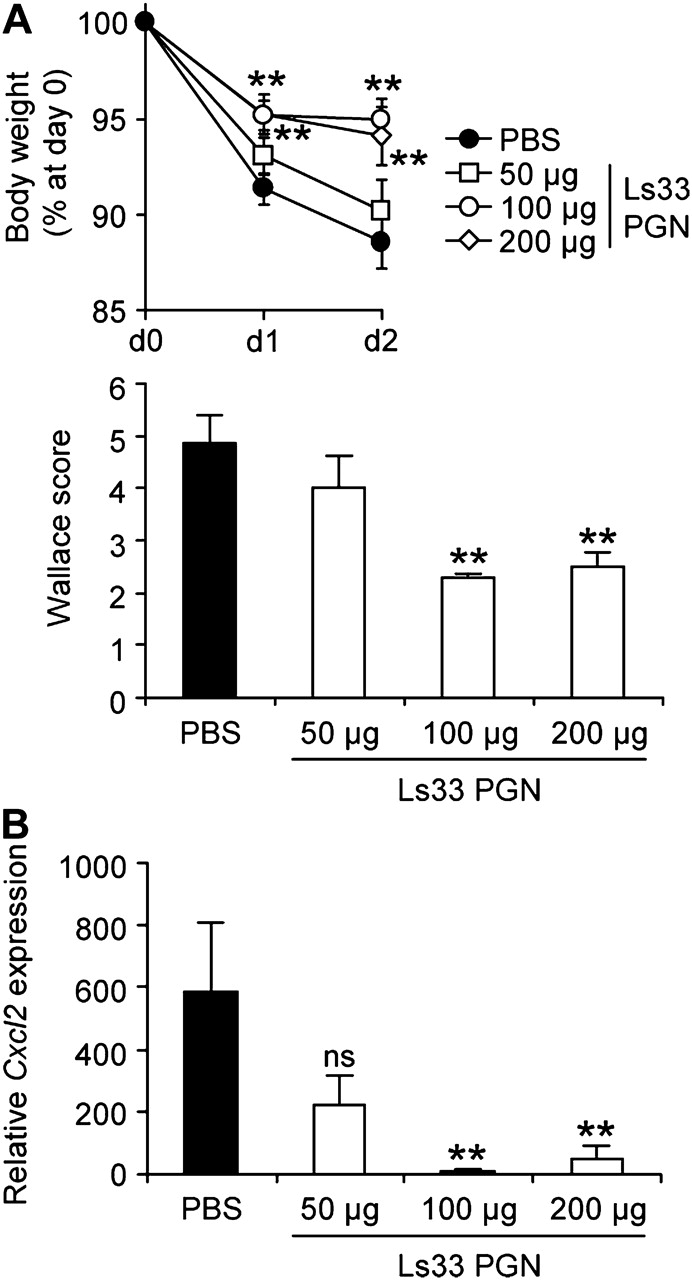
Oral administration of Ls33 peptidoglycan (PGN) rescues BALB/c mice from trinitrobenzene sulfonic acid (TNBS)-induced colitis. BALB/c mice were fed for five consecutive days with phosphate-buffered saline (PBS) or varying concentrations of Ls33 PGN (50, 100 or 200 μg) before administration of TNBS. (A) Loss of body weight and Wallace score and (B) quantification of colonic Cxcl2 gene expression were evaluated 2 days after induction of colitis. Data represent the mean of 8–10 mice per group. Error bars represent the SEM (ns, not significant; **p<0.01).
The protective effect of Lactobacillus PGN against TNBS-induced colitis is strain specific
Since lactobacilli have strain-specific anti-inflammatory properties,15 we investigated the potential role of PGN in this strain specificity. Hence, we compared the protective capacity of various concentrations of Ls33 PGN with PGN purified from L. acidophilus NCFM, a strain we previously showed to be non-protective in the TNBS colitis model. While intraperitoneal administration of Ls33 PGN reduced the macroscopic inflammation score in a dose-dependent manner, NCFM PGN had no protective effect on colitis (figure 4A). These findings were corroborated by histological analysis (figure 4B). Taken together, these data suggest that the immunomodulatory properties of PGNs are correlated with the anti-inflammatory capacity of the strain from which they derive.

Peptidoglycan (PGN) from selected strains rescues mice from trinitrobenzene sulfonic acid (TNBS)-induced colitis. BALB/c mice were injected intraperitoneally with phosphate-buffered saline (PBS; n=10), 50 μg (n=5) or 200 μg (n=10) of PGN from Ls33 (Ls33 PGN) or NCFM (NCFM PGN) before induction of colitis. (A) Wallace score evaluated 2 days after induction of colitis. (B) Ameho scores illustrated with representative sections of the distal colon of healthy mice or mice with TNBS-induced colitis, injected intraperitoneally with PBS or 200 μg of PGNs (original magnification, ×20). Data represent mean values with the SEM (*p<0.05; **p<0.01; ***p<0.001 vs PBS-treated mice).
Ls33 PGN induces regulatory DCs in vitro able to rescue mice from colitis after adoptive transfer
We previously reported that Ls33 could induce semi-mature BMDCs in vitro, able to protect mice from colitis after adoptive transfer, while NCFM did not.16 Therefore, we compared the ability of PGN from Ls33 and NCFM to activate BMDCs derived from BALB/c mice in vitro. As expected, LPS fully matured BMDCs, with enhanced surface expression of CD40, CD80 and CD86, as compared with untreated BMDCs. While NCFM PGN strongly upregulated levels of these co-stimulatory markers, Ls33 PGN induced only partial maturation of BMDCs (figure 5A). However, in contrast to LPS, both PGNs were found to be poor inducers of proinflammatory mediators such as IL-12p70 or CCL3 (figure 5B). Interestingly, only Ls33 PGN induced high amounts of IL-10 (figure 5B). In vivo, while intraperitoneal injection of untreated or NCFM PGN-treated BMDCs did not have any effect on colitis, adoptive transfer of Ls33 PGN-treated BMDCs induced a significant reduction of colitis (p<0.001) (figure 5C). Using BMDCs derived from C57BL/6 WT and Nod2−/− mice, we confirmed the induction of regulatory DCs in this genetic background and also demonstrated the NOD2 dependency (figure 5D).

Ls33 peptidoglycan (PGN) specifically induces regulatory dendritic cells (DCs) in vitro which protect mice from colitis after adoptive transfer. (A and B) Bone barrow-derived DCs (BMDCs) generated from BALB/c mice were stimulated overnight with Ls33 PGN (10 μg/ml), NCFM PGN (10 μg/ml), lipopolysaccharide (LPS; 100 ng/ml) or were left untreated. (A) Co-stimulatory molecules expressed on BMDCs were analysed by gating on CD11c-positive cells by flow cytometry. Histograms show CD40, CD80 and CD86 expression profiles of unstimulated (grey histograms) and PGN- or LPS-treated (dark grey histograms) BMDCs. Thin lines represent isotype controls from a pool of untreated and stimulated BMDCs. The data are representative of three independent experiments. (B) Cytokines and chemokine levels were determined in culture supernatant by ELISA. Data represent the mean with the SEM of five independent experiments performed in duplicate (**p<0.01). (C) BALB/c mice were intraperitoneally injected with unstimulated (DC) Ls33 PGN (Ls33 PGN DC)- or NCFM PGN (NCFM PGN DC)-treated BMDCs concomitantly with trinitrobenzene sulfonic acid (TNBS) instillation. The Wallace score was determined 2 days later. Data represent mean values of 10 mice per group with the SEM (***p<0.001 vs phosphate-buffered saline (PBS)-treated mice). (D) Groups of 10 wild-type (WT) C57BL/6 mice were intraperitoneally injected with BMDCs derived from WT or Nod2−/− C57BL/6 mice previously stimulated or not (DC) in vitro with Ls33 (multiplicity of infection of 10:1) (Ls33 DC) or 10 μg/ml Ls33 PGN (Ls33 PGN DC) just before TNBS instillation. The Wallace score was determined 3 days later. Error bars represent the SEM (*p<0.05; **p<0.01 vs PBS-treated mice).
Ls33 PGN harbours additional muropeptides including M-tri-Lys, a NOD2 ligand displaying anti-inflammatory properties
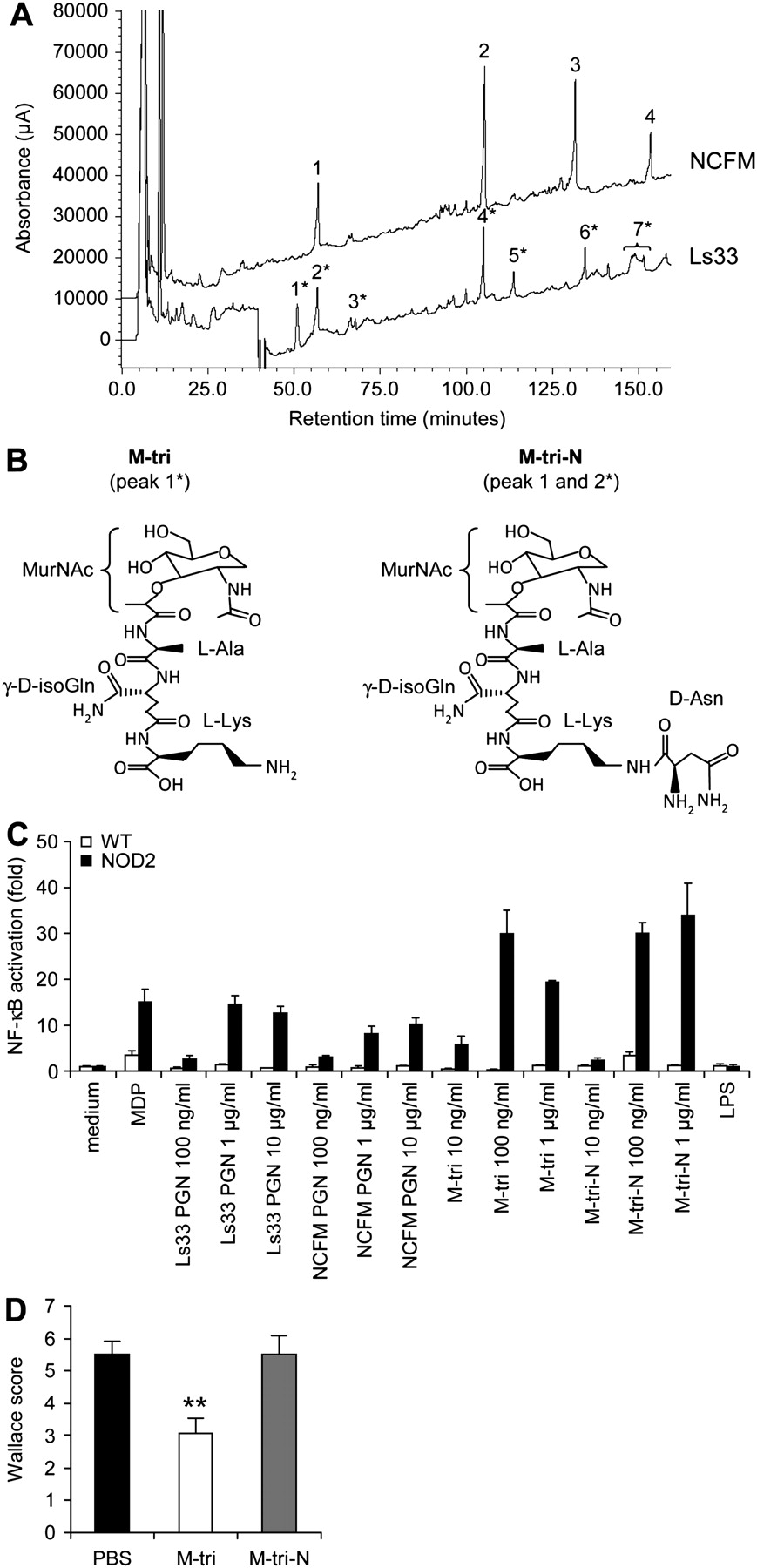
To determine the underlying molecular basis of the strain-specific anti-inflammatory effect of Ls33 PGN, we analysed the structure of both Ls33 and NCFM PGN and compared the resulting muropeptides obtained after mutanolysin (muramidase) digestion. We observed that strain Ls33 produced muropeptides that were not released by strain NCFM (figure 6A). As described in the literature, both strains contain the Lys-d-Asn-type PGN, with an epsilon-link between l-Lys and d-Asn. As reported in table 1, both PGNs carried monomers, dimers and trimers of GlcNAc-MurNAc-l-Ala-γ-d-iGln-l-Lys-(ε-d-Asn) (GM-tri-Lys-d-Asn, peak 1 and 2* for NCFM and Ls33 PGN, respectively). However, while NCFM produced exclusively muropeptides carrying the epsilon-linked d-Asn (GM-tri-N), Ls33 produced in addition significant amounts (∼10% of the total amount of muropeptides released) of the monomer without the d-Asn, namely the GM-tri-Lys (GlcN-MurNAc-l-Ala-γ-d-isoGln-l-Lys, GM-tri, peak 1*). Both chemically synthesised muropeptides (core structure indicated in figure 6B), as well as purified PGNs, were sensed by NOD2 (figure 6C). However, intraperitoneal administration of both muropeptides revealed that only M-tri-Lys was able to rescue BALB/c mice from TNBS-induced colitis (figure 6D). Its protection was dose-dependent and correlated with a reduction of colonic proinflammatory gene expression (Supplementary figure 5). Moreover, M-tri-Lys was also able to rescue C57BL/6 WT mice but not their Nod2−/− counterparts, indicating that its protective effect is NOD2-dependent (figure 7A). Furthermore, while the protective effect of Ls33 PGN is lost in MyD88-deficient mice, the protective effect of M-tri-Lys is MyD88-independent (figure 7B). Neither Ls33 nor NCFM produced detectable amounts of the muropeptide GlcNAc-MurNAc-l-Ala-d-Glx (GMDP), which comprises the minimal structure of PGN detected by NOD2 (MDP). However, Ls33 accumulated additional muropeptides (peaks 5* and 7* in figure 6), corresponding to muropeptides partially digested by endogenous PGN hydrolases. In particular, peak 7* is a mixture of two muropeptides that were cleaved simultaneously by an amidase and by γ-d-glutaminyl-lysine endopeptidase.

Structural analysis of Ls33 and NCFM peptidoglycans (PGNs) reveals additional muropeptides in the Ls33 strain including a NOD2 ligand exhibiting anti-inflammatory capacities. (A) The composition of Ls33 and NCFM muropeptides was analysed by reversed-phase high-performance liquid chromatography (HPLC) separation. Ls33 muropeptides are marked by an asterisk (see table 1). (B) Core chemical structure of the M-tri and the M-tri-N (peak 1* and peaks 1 and 2*, respectively). (C) Wild-type (WT) and murine NOD2-expressing HEK293 cell lines transfected with the nuclear factor-κB (NF-κB)-inducible luciferase reporter plasmid were stimulated with muramyl dipeptide (MDP; 1 μg/ml), lipopolysaccharide (LPS; 100 ng/ml) or increasing concentrations of PGNs (100 ng/ml, 1 μg/ml and 10 μg/ml) or muramyl peptides (10 ng/ml, 100 ng/ml and 1 μg/ml) for 24 h before the quantification of the NF-κB-dependent luciferase activation. Data represent mean values of triplicate cultures with the SD. The results are expressed in fold NF-κB activation reported to the level of unstimulated cells. (D) Macroscopic score of BALB/c mice injected intraperitoneally with phosphate-buffered saline (PBS) or 20 μg of synthetic muropeptides before induction of colitis. Data represent mean values of 10 mice with the SEM (**p<0.01 vs PBS-treated mice).
Structures and molecular masses of Ls33 and NCFM muropeptides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The anti-inflammatory property of M-tri-Lys is NOD2-dependent but MyD88-independent. (A, B) Wild-type (WT), Nod2−/− or MyD88−/− C57BL/6 mice were injected intraperitoneally with phosphate-buffered saline (PBS) or 20 μg of M-tri concomitantly with trinitrobenzene sulfonic acid (TNBS) instillation. The Wallace score was determined 3 days later. Data represent mean values of 8–9 animals with the SEM (***p<0.001 vs PBS-treated mice).
Discussion
It remains extremely important to unravel the mechanism of action of probiotics, notably to understand their strain-specific effect in IBD. In this study, we demonstrated that PGN can promote anti-inflammatory properties upon sensing of NOD2. We observed that the protective effect of Ls33 administration was correlated with an upregulation of IL-10 production in the colon, an effect which was abrogated in NOD2-deficient mice. Since NOD2 is a member of the NLR family involved in PGN sensing,17 we evaluated the role of PGN in the anti-inflammatory capacity of the strain Ls33. We showed that a single systemic administration of Ls33 PGN rescued mice from colitis, an effect associated with a downregulation of key inflammatory effectors (TNFα, IL-1β and IL-6) and an upregulation of genes involved in the IDO immunosuppressive pathway in the colon. Although a decrease in Il10 gene expression was observed in Ls33 PGN-treated mice, a local increase of the IL-10 protein was positively correlated with the observed protection by the PGN. This apparent discrepancy should be linked to a difference in the kinetics of Il10 gene expression, which is probably faster in PGN-treated mice. The role of IL-10 was however confirmed, since the protective effect of PGN was abrogated in Il-10-deficient mice. Treg cells are known to play a critical role in the control of inflammatory responses, notably CD4+CD25+FoxP3+ Treg cells are central to the maintenance of immunological homeostasis and tolerance.27 Recent observations have shown that Treg cells can be markedly promoted by a specialised population of mucosal CD103+ DCs via a TGFβ- and RA-dependent mechanism,3 which abrogates their tolerogenic properties in colitic mice.28 Interestingly, we could correlate the protective effect of Ls33 PGN to the increase in both CD4+FoxP3+ Treg cells and CD103+ DCs in MLNs of PGN-treated mice. Moreover, we also observed an increase of both Ifng and Indo mRNA levels in MLNs enriched with CD11c+ DCs. No enhancement of Ifng was found in CD4+ T cells (data not shown), suggesting that its expression was not linked to Th1 expansion. In addition to the well-described proinflammatory properties typically associated with Th1-mediated inflammation, increased IFNγ expression in the setting of TNBS colitis has also been reported to exhibit protective anti-inflammatory properties. Moreover, IFNγ is the main inducer of the IDO-mediated immunosuppressive pathway.29 The IDO-dependent pathway was recently shown to be important for CD103+ intestinal DCs as it preferentially drives the CD4+FoxP3+ Treg cell expansion.30 Therefore, our results emphasise that Ls33 PGN favours the induction of tolerogenic responses. This is supported by the fact that this PGN was also able to induce in vitro IL-10-producing partially mature DCs, able to rescue mice from colitis upon adoptive transfer. This protective effect was completely abolished when DCs were derived from Nod2-deficient mice, supporting the hypothesis that the ‘tolerogenic’ effect of Ls33 PGN is mediated through the induction of regulatory DCs via NOD2 signalling. More interestingly, the observed anti-inflammatory properties were strain specific, since they were not obtained with PGN derived from the non-anti-inflammatory strain L. acidophilus NCFM. While most research has highlighted the role of PGN in the pathogenesis of various harmful bacteria,31 few papers actually reported beneficial effects of this bacterial compound,32–34 emphasising mainly its immune-potentiating role. Very recently, Shida et al35 showed that PGN from selected lactobacilli was able to inhibit IL-12 production by macrophages. Our findings reveal that PGN from selected lactobacilli can indeed promote functional anti-inflammatory activity.
NOD2 sensing with MDP was first shown to enhance proinflammatory pathways.36 37 Since genetic variation in Nod2/Card15 is associated with susceptibility to CD,18 it has been suggested that this can lead to defects in sensing bacteria and to abnormal T cell-mediated responses, resulting in tissue inflammation.38 Watanabe et al39 also showed that MDP activation of NOD2 can negatively regulate TLR2 responses and diminish Th1 responses. The authors demonstrated that synthetic MDP is able to protect mice from colitis in a NOD2-dependent manner.40 These findings further support the hypothesis that NOD2 signalling can downregulate inflammation and that mutations in Nod2 can contribute to active IBD by leading to excessive TLR2 cytokine responses.
We also demonstrated that the Ls33 PGN protective effect could be achieved by the most convenient oral route. Importantly, the active dose of 100–200 μg of PGN per mice per day corresponded to the quantity of PGN derived from a classical dose of 108 CFU of probiotic lactobacilli administrated daily to mice. This effect was specific to PGN since we clearly demonstrated that (1) the PGN preparation was devoid of TLR2 contaminants such as LTA and (2) Ls33 LTA administration by itself did not exhibit anti-inflammatory potential. Moreover, since the protective effect of Lactobacillus PGN depends of the strain, we hypothesised that a specific PGN structures might be the driving force behind the selective anti-inflammatory properties of selected probiotic strains.
PGN, the major component of the Gram-positive cell wall, is composed of a network of glycan strands formed by alternating residues of β-(1,4)-linked N-acetylglucosamine (GlcNAc) and N-acetylmuramic acid (MurNAc) cross-linked by short peptides. In most lactobacilli, this stem peptide is composed of the pentapeptide chain l-Ala-γ-d-Glx-l-Lys-d-Ala-d-Ala. Since the two strains compared in this paper share the same l-Lys-d-Asn cross-bridge (A4 α subtype), we hypothesised that the differences in anti-inflammatory potential observed are linked to a difference in the turnover of the PGN, affecting the release of biological active muropeptides. Indeed, structural analysis demonstrated that only strain Ls33 incorporated the muropeptide GlcNAc-MurNAc-l-Ala-γ-d-iGln-l-Lys (M-tri-Lys), which was completely absent in the NCFM PGN. Moreover we highlighted that the anti-inflammatory potential of M-tri-Lys, described before as a NOD2 ligand,41 was indeed NOD2- dependent. Surprisingly, while we also demonstrated that NOD2 sensed both Ls33 and NCFM PGN as well as M-tri-N, neither NCFM PGN nor M-tri-N exhibited a protective effect. The opposite effect of both muropeptides could be linked to a differential transport. The PepT1 di/tripeptide transporter has been recently shown to be involved in the anti-inflammatory effect of the tripeptide KPV.42 Interestingly, this transporter, normally expressed in the small intestine, is induced in chronically inflamed colon and its transport activity has been proposed as a potential new target for anti-inflammatory treatments. Moreover, it has been recently shown that the delivery of NOD2 ligands following internalisation and processing of bacteria in phagolysosomes appeared also to be mediated by dedicated transporters such as PEPT1 or PEPT2.43 We can therefore speculate that only the tripeptide M-tri-Lys released from Ls33 PGN is correctly delivered to the cytosolic compartment, allowing NOD2 sensing. Furthermore, while the sensing of PGN by TLR2 still remains controversial, it was recently shown that PGN translocation could be mediated by forming a complex with a PGN recognition protein (PGLYRP-3) and the subsequent TLR2-mediated phagocytosis pathway.44 Phagosome maturation seems to be controlled by TLR signals through the adaptator MyD88.45 This could explain the MyD88 dependency of the protective effect of Ls33 PGN, while the effect of M-tri-Lys is strictly NOD2-dependent.
Our results indicated that Ls33 seems to have a very active PGN turnover, in contrast to NCFM. This could promote the accumulation and release of the M-tri-Lys precursor and as such explain the protective effect of Ls33 PGN. Even if both PGNs can be sensed by NOD2 in a transfected HEK cell line, it remains to be evaluated if such molecules can differentially reach NOD2 under physiological conditions. PGN recognition proteins (PGRPs) are innate immunity proteins that can exhibit antimicrobial, proinflammatory, but also anti-inflammatory, responses,46 notably through their hydrolytic activities. We could hypothesise that the PGN differential recognition or degradation process could lead to such differential anti-inflammatory effects of Ls33 and NCFM PGNs.
Considering the results obtained in this study, it might be rewarding to investigate in more detail the structure and turnover of PGNs from other LAB. The production of isogenic pairs of mutants may yield important information and provide useful hints for the selection of the most suitable strains for future therapeutic or prophylactic applications, in particular for the treatment of IBD. Moreover, this work points out the potential use of purified PGN or specific muropeptide derivatives as new immune intervention tools.
Acknowledgments
We thank B. Foligné and V. Peucelle for technical assistance. We are very grateful to Danisco for supplying the bacterial strains. We warmly thank J.-C. Sirard for scientific advice and for the generous gift of MyD88−/− mice, and M. Simonet for fruitful discussions.
References
Supplementary materials
Web Only Data for gut.2010.232918
Files in this Data Supplement:
Footnotes
See Commentary, p 1026
Linked articles 241026.
Funding This work was financially supported by l'Institut Pasteur de Lille, l'Association François Aupetit, by INSERM and by a European Research Council starting grant (202283-PGNfromSHAPEtoVIR) to IGB. EMF was supported by a doctoral fellowship from the Ministère de l'Enseignement Supérieur et de la Recherche.
Competing interests None.
Ethics approval Animal experiments were performed in compliance with European guidelines in accredited establishments (no. A59107; Institut Pasteur de Lille and no. B 75 15-01 Institut Pasteur, Paris). All animal protocols were approved by the locally appointed investigational review board (number 86/609/EEC).
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Commentary
- PostScript