Article Text

Abstract
Objectives During pancreatitis, specific transcriptional programmes govern functional regeneration after injury. The objective of this study was to analyse the dynamic regulation of pancreatic genes and the role of transcriptional regulators during recovery from pancreatitis.
Design Wild-type and genetically modified mice (Hnf1α−/− and Ptf1a+/−) were used. After caerulein or l-arginine induced pancreatitis, blood or pancreata were processed for enzymatic assays, ELISA, histology, immunohistochemistry, western blotting and quantitative reverse transcriptase-PCR. Nr5a2 promoter reporter and chromatin immunoprecipitation assays for Hnf1α were also performed.
Results After caerulein pancreatic injury, expression of acinar and endocrine genes rapidly decreased, but eventually recovered, depicting distinct cell-type-specific patterns. Pdx1 and Hnf1α mRNAs underwent marked downregulation, matching endocrine/exocrine gene expression profiles. Ptf1a, Pdx1 and Hnf1α protein levels were also reduced and recovered gradually. These changes were associated with transient impairment of exocrine and endocrine function, including abnormal glucose tolerance. On l-arginine pancreatitis, changes in Ptf1a, Pdx1 and Hnf1α gene and protein expression were recapitulated. Reduced Hnf1α and Ptf1a levels after pancreatitis coincided with increased acinar cell proliferation, both in Hnf1α−/− and Ptf1a+/− mice. Moreover, Hnf1α−/− mice had reduced Ptf1a protein as well as transcripts for Ptf1a and digestive enzymes. Dispersed acini from Hnf1α−/− mice showed suboptimal secretory responses to caerulein. Bioinformatics analysis did not support a role for Hnf1α as a direct regulator of digestive enzyme genes. Instead, it was found that Hnf1α binds to, and regulates, the promoter of Nr5a2, coding an orphan nuclear receptor that regulates acinar gene expression.
Conclusions Dynamic changes in gene expression occur on pancreatitis induction, determining altered exocrine and endocrine function. This analysis uncovers roles for Hnf1α in the regulation of acinar cell determination and function. This effect may be mediated, in part, through direct regulation of Nr5a2.
- Pancreatitis
- Ptf1a
- Hnf1α
- Nr5a2
- pancreatic fibrosis
- pancreatic pathology
- pancreatic cancer
- cystic fibrosis
- pancreas
- pancreatic cancer
- pancreatitis
- cancer
- carcinogenesis
- pancreatic fibrosis
- pancreatic physiology
- diabetes mellitus
- ageing
- αß t cells
- gene expression
- molecular carcinogenesis
- mucins
- epithelial differentiation
- molecular oncology
- pancreatic tumours
Statistics from Altmetric.com
- Pancreatitis
- Ptf1a
- Hnf1α
- Nr5a2
- pancreatic fibrosis
- pancreatic pathology
- pancreatic cancer
- cystic fibrosis
- pancreas
- pancreatic cancer
- pancreatitis
- cancer
- carcinogenesis
- pancreatic fibrosis
- pancreatic physiology
- diabetes mellitus
- ageing
- αß t cells
- gene expression
- molecular carcinogenesis
- mucins
- epithelial differentiation
- molecular oncology
- pancreatic tumours
Significance of this study
What is already known about this subject?
Regeneration of the pancreas proceeds to completion after a single bout of caerulein-induced pancreatitis.
Functional pancreatic regeneration after injury is governed by specific transcriptional programmes that recapitulate, in part, embryological programmes.
A specific role for Hnf1α in exocrine pancreas terminal differentiation and function has not yet been proposed.
What are the new findings?
We provide evidence for injury-responsive regulated expression of Ptf1a, Pdx1 and Hnf1α in acinar and endocrine cells. Importantly, we also provide evidence for transient deranged expression of genes under the control of these transcriptional regulators, resulting in inappropriate pancreatic function.
Hnf1α is regulated in acute pancreatitis during initiation and recovery stages at the mRNA and protein levels.
In Hnf1α−/− mice, the concentrations of transcripts for digestive enzymes and Ptf1a are reduced, and acinar cell secretory function is suboptimal.
Hnf1α binds and regulates the promoter of Nr5a2, a regulator of some of the genes coding for digestive enzymes.
Significance of this study
How might it impact on clinical practice in the foreseeable future?
Our findings provide experimental evidence for highly regulated fluctuations in transcriptional regulators after pancreatitis, which result in transient endocrine and exocrine insufficiency preceding full recovery. Impaired or absent activity of these proteins may result in permanent derangement of pancreatic physiology after injury.
Knowledge of the transcriptional control of pancreatic cell dynamics in response to injury is key to understanding the events involved in pancreatic regeneration, the development of chronic fibrogenic disease, and pancreatic carcinogenesis. Full characterisation of transcriptional networks that govern pancreatic functional regeneration opens the study of molecular pathways that may be subject to positive or negative modulation.
Our findings contribute insights into pancreatic pathophysiology that may be valuable for understanding events during acute and chronic pancreatitis and perhaps during the steps that herald the initiation of carcinogenesis.
Introduction
Pancreas formation, lineage specification and terminal differentiation of exocrine and endocrine cells depend on the activity of a highly regulated transcriptional factor code.1 Pdx1 and Ptf1a are expressed in pancreatic progenitors and are required for pancreas formation and activation of expression of insulin and digestive enzyme genes, respectively.2 Mice lacking either of them display severe defects in early pancreas formation.2 ,3 Mice in which Ptf1a expression levels are reduced by >50% display pancreatic hypoplasia, reduced ß cell number, and glucose intolerance.4 In the adult, Ptf1a is exclusively expressed in acinar cells where, together with a ubiquitous E-protein and recombination signal-binding protein J-like (Rbpjl), it constitutes the heterotrimeric complex, pancreas transcription factor 1 (PTF1).5 During early pancreatic development, Rbpj is the Rbp isoform partner in PTF1 and is required for pancreatic growth and morphogenesis.6 Beginning at the secondary transition, Rbpj is replaced by Rbpjl in PTF1 and, in the adult pancreas, only Rbpjl participates in this complex.5 PTF1 binds and activates the promoter of genes encoding the secretory digestive enzymes.5
Pdx1 is essential for the commitment of the foregut endoderm to common pancreatic precursors, where it regulates the expression of a large set of target genes; lack of Pdx1 is associated with pancreatic agenesis.7–9 Later on, Pdx1 participates in both endocrine and exocrine differentiation; its levels are upregulated in ß cells and downregulated in acinar and ductal cells. Forced overexpression of Pdx1 in differentiating acinar cells leads to ductal metaplasia.10 In adult mice, Pdx1 is mainly expressed in ß cells, where it controls the expression of endocrine genes, including insulin. Pdx1+/− mice show defects in glucose homoeostasis, and conditional repression of Pdx1 in adult mice leads to impaired insulin expression.11 ,12
Several other transcription factors participate in gene regulatory networks in the pancreas. In adult mice, Hnf1α is constitutively expressed in both acinar and endocrine cells.13 One function attributed to Hnf1α in the liver and pancreas is the maintenance of differentiated cell phenotypes by modulating broad cell-specific genetic programmes.14 ,15 In human islets, Hnf1α binds directly to >100 target genes, eight of them encoding transcription factors, and Hnf1α deficiency in mouse islets leads to altered expression of hundreds of genes.14 ,15 Hnf1α null mice are viable and develop early diabetes, fatty liver and renal dysfunction.16 In addition, HNF1α mutations are the cause of type 3 maturity-onset diabetes of the young (MODY3).17 Intriguingly, patients with MODY3 present lower levels of faecal elastase 1 than control subjects, although, in general, elastase concentration falls within the normal range.18 Despite this observation, a role for Hnf1α in exocrine pancreas differentiation and function has not yet been proposed.
Mild acute pancreatitis can be induced in rodents by repeated caerulein administration. This approach yields an acute inflammatory reaction of the pancreas that resolves completely in 14 days. This model has allowed the identification of important signalling pathways involved in recovery from pancreatic damage, such as Notch and Hedgehog.19–21 Here, we first analyse the dynamic regulation of acinar, endocrine and ductal genes—as well as pancreatic transcription factors—after the induction of acute caerulein pancreatitis in mice. We find that—in addition to Ptf1a and Pdx1—Hnf1α is also regulated. Hnf1α−/− mice display increased acinar cell proliferation in basal conditions and after pancreatitis induction. Furthermore, they display reduced expression of digestive enzyme transcripts and suboptimal enzyme secretion in vitro. Hnf1-binding motif sequences are not enriched in these genes. In acinar cells, Hnf1α binds to, and regulates, the promoter of Nr5a2, which codes for an orphan nuclear receptor that is also modulated during pancreatitis and participates in expression of genes coding for digestive enzymes.22
Materials and methods
Detailed materials and methods are given in the online supplementary data.
Mice
Male C57BL/6J mice weighing 28–32 g were used. Hnf1α−/− mice in a C57BL/6J background were bred as previously described.23 Ptf1aCre/+ knock-in mice were obtained from C Wright (Vanderbilt University, Nashville, Tennessee, USA).2 Control animals were sex- and aged-matched littermates. For detailed procedures on induction of acute pancreatitis, glucose tolerance test, sampling, amylase and insulin assays, ultrastructural analysis, and preparation of isolated acini for amylase secretion, see the online supplementary data. Procedures were approved by the Animal Experimentation Committee of Hospital Vall d'Hebron and Instituto de Salud Carlos III.
Immunohistochemistry and western blotting
Pancreata were formalin-fixed and paraffin-embedded for immunohistochemistry. For western blotting, total pancreatic homogenates were prepared in radioimmunoprecipitation buffer with protease inhibitors.
Analysis of mRNA expression
Quantitative reverse transcriptase (qRT)-PCR expression was carried out using either TaqMan chemistry or SYBR-Green dye; data were expressed relative to expression of Tbp or Hprt. A list of primers used is provided in online supplementary table 1. The threshold cycle (Ct) methodology was used to calculate relative quantities of mRNA products.24 ,25 All values were expressed as the mean of the relative quantities of mRNA relative to normal (basal) pancreas set at 1.0. In general, measurements were performed in triplicate; all major findings were replicated in independent assays.
Hnf1α chromatin immunoprecipitation (ChIP)
To demonstrate binding of Hnf1α to the Nr5a2 promoter, ChIP using Hnf1α antibodies was performed as described in online supplementary materials and methods.14 ,15
Statistical analysis
All data are expressed as means±SEM. Differences between means were analysed by Student t test or analysis of variance followed by Dunnett's multiple comparison test, depending on the number of the groups analysed. Statistically significant difference was defined as a p value <0.05. All data analyses were performed using the GraphPad Prism statistical program (Graph Pad Prism 4.0a for Macintosh).
Results
Caerulein administration resulted in oedema, inflammatory cell infiltration and some acinar cell collapse. These changes were detected at 7 h and peaked at 24 h. As expected, an acute increase in serum amylase was observed 7 h after the first caerulein injection (9768±3834 U/l vs basal values of 1855±569; p<0.05, n=5). By day 14 morphological and biochemical recovery was complete.
Acute pancreatic injury induces distinct changes in gene expression in different pancreatic cell types
To analyse changes in gene expression, we used qRT-PCR to examine acinar, endocrine and ductal genes (figure 1 and online supplementary table 2). Transcripts coding for digestive enzymes showed a consistent pattern, characterised by a rapid decrease in mRNA levels followed by a slow recovery that was still incomplete by day 4. In response to caerulein, all the endocrine genes examined also showed a consistent, but distinct, pattern characterised by a rapid decrease—with a nadir at 24 h—and complete or almost complete recovery by 48 h. Transcript levels of well-characterised ductal genes (Krt19, Krt7, Cftr and Ceacam1) showed a distinct, though more diverse, pattern characterised by an early increase followed by subsequent normalisation (figure 1). Levels of desmin and α-Sma transcripts, two stellate cell markers, were analysed: a modest increase in desmin mRNA was observed at days 5 and 8 (online supplementary figure 1). Expression patterns of transcription factors involved in pancreatic cell differentiation varied substantially. Despite profound and sustained reduction in acinar gene expression, Ptf1a transcript levels did not show consistent changes. By contrast, Rbpjl, Pdx1 and Hnf1α mRNAs underwent marked reduction over time, depicting patterns matching exocrine or endocrine gene expression profiles. Rbpjl mRNA levels showed a sharp downregulation followed by a slow recovery, similar to the pattern observed for acinar genes. Pdx1 mRNA closely followed the endocrine gene expression pattern. Hnf1α mRNA expression displayed a specific profile, characterised by an initial discrete upregulation followed by a more profound and sustained downregulation.
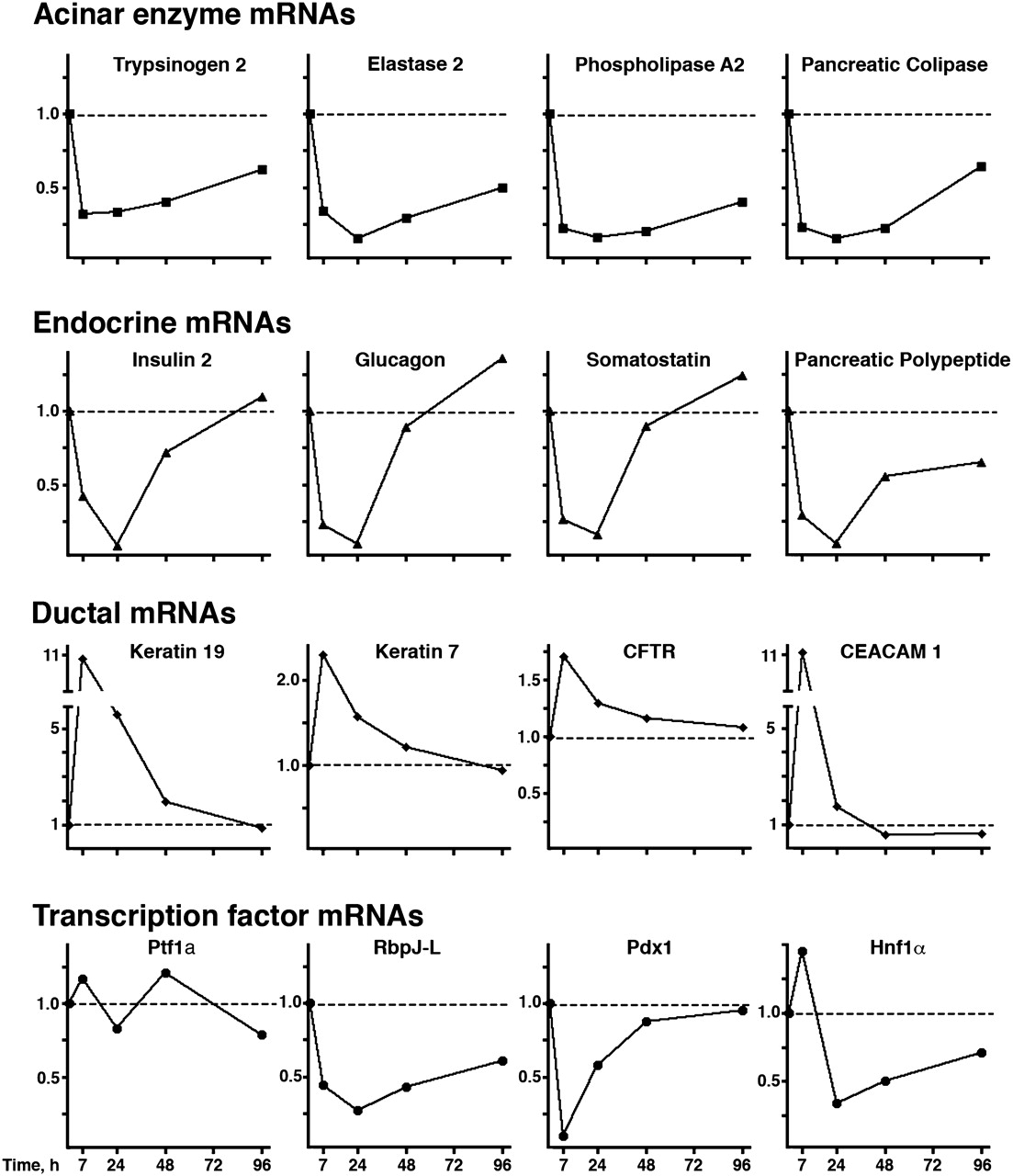
Quantitative reverse transcriptase-PCR analysis of expression of selected genes after induction of caerulein pancreatitis. Dotted lines refer to levels in samples at time 0. Expression of acinar enzyme mRNAs is markedly reduced, depicting a similar pattern for all genes examined. Expression of endocrine gene transcripts is sharply reduced for a short time after pancreatitis, outlining a distinct common pattern. Ductal gene transcripts show an early upregulation with subsequent reduction to baseline levels. Dynamic patterns of expression in selected pancreatic transcription factor transcripts are individually distinct. (y axis: fold over basal, RQ)
These findings indicate that acinar, endocrine and ductal genes experience distinct changes in expression during acute pancreatitis, and that transcription factors involved in pancreatic cell differentiation probably contribute to these changes.
Dynamic changes in the expression of Ptf1a, Pdx1 and Hnf1α in response to pancreatic injury
We analysed the expression of Ptf1a, Pdx1 and Hnf1α during acute pancreatitis induction and recovery using western blotting and immunohistochemistry (figure 2). Early after induction, the level of all three proteins was greatly reduced. Nuclear Ptf1a expression in acinar cells was sharply reduced 3 h after initiation of pancreatitis and recovered subsequently (figure 2A). These findings are similar (although at a different time scale) to those reported in rats.26 In normal pancreas, Pdx1 was strongly detected in the nucleus of β cells and weakly in acinar cells. Pdx1 was markedly downregulated 7 and 24 h after pancreatitis induction (figure 2B). A slight cytoplasmic immunoreactivity persisted in some endocrine cells. Pdx1 nuclear expression in β cells had recovered at 48 h (figure 2B). Hnf1α was strongly expressed in both islet and acinar cells and was expressed at low or undetectable levels in ductal cells; it became almost undetectable 24 h after the initiation of pancreatitis, and had partially recovered at day 4 (figure 2C).
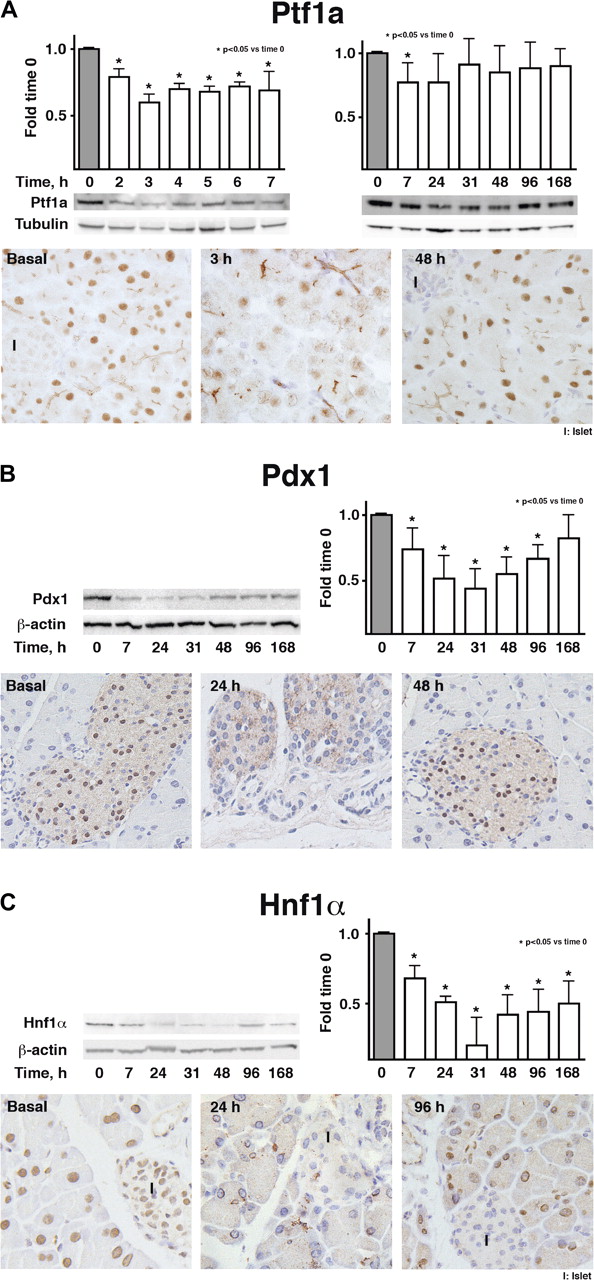
Transcription factor protein expression in pancreatic tissue after pancreatitis induction. Columns represent arbitrary units calculated by densitometry of western blots and normalised using tubulin or ß-actin (n=4). Representative immunoperoxidase staining is shown below. Original magnification ×400.
To assess whether similar changes in transcription factor expression occurred in other models of experimental acute pancreatitis, we used intraperitoneal l-arginine administration.27 Extensive acute pancreatic damage occurred, and mortality over 7 days was 41%. Tissue damage was variable, but clearly detectable at day 2, peaked at days 3–4, and evolved long beyond day 10. Serum amylase increased at day 2 (online supplementary figure 2A) and then returned to basal levels.
On western blotting, expression of Ptf1a, Pdx1 and Hnf1α was found to decrease in response to l-arginine with variable degrees of impairment (online supplementary figure 2B). Hnf1α and Ptf1a showed a consistent pattern: reduced levels were detected as early as 7 h–1 day (Ptf1a) or days 1–2 (Hnf1α), when very little tissue damage was present; a greater reduction occurred at 2–4 days, after which expression recovered. Unlike in caerulein pancreatitis, Pdx1 expression was only moderately reduced at days 2–3 and recovered thereafter.
On immunohistochemistry, Hnf1α, Ptf1a and Pdx1 expression mirrored western blotting findings (online supplementary figure 3). Hnf1α and Ptf1a were undetectable in a high proportion of normal-appearing acinar cells at days 2–3 (online supplementary figure 3A,B) and slowly, but incompletely, recovered at days 7–10. Ptf1a and Hnf1α expression in acinar cells displayed a mosaic distribution. Unlike in caerulein-induced pancreatitis, islet Hnf1α expression was marginally affected and only at early time points (online supplementary figure 3A). Remarkably, from days 2–4, Ptf1a showed mainly a cytoplasmic distribution (online supplementary figure 3B, inset). We conclude that Ptf1a and Hnf1α are extensively regulated in the two models of acute pancreatitis analysed.
Impaired transient expression of Ptf1a, Pdx1 and Hnf1α in response to acute pancreatitis is associated with abnormal transient exocrine and endocrine function
These findings suggest that, in caerulein-induced pancreatitis, both exocrine and endocrine function might be affected as a result of transcription factor downregulation. Indeed, amylase mRNA and pancreatic activity were greatly reduced 4 h after pancreatitis induction, at a time when Ptf1a expression was severely compromised (figure 3A,B). The reduction in amylase activity cannot be fully explained by the loss of acinar cell mass associated with pancreatitis. Importantly, amylase activity increased rapidly (figure 3B) after Ptf1a recovery.

Exocrine and endocrine function at the indicated times after caerulein pancreatitis induction. (A) Amylase mRNA levels in pancreas (quantitative reverse transcriptase (qRT)-PCR). (B) Amylase activity in the pancreas. (C) Insulin 2 mRNA levels in total pancreas (qRT-PCR). (D) Plasma insulin concentration (ELISA). (E) Glycaemia in normal fasting mice and 20 min after glucose overload (GTT); basal conditions (grey bars); 24 h after caerulein pancreatitis (white bars).
In agreement with the reduced expression of Pdx1 and Hnf1α proteins at 24 h, insulin 2 pre-mRNA levels were also reduced (figure 3C), as was fasting insulinaemia (figure 3D). Fasting glycaemia was unaffected, but the response to glucose overload was abnormal (figure 3E).
Reduced Hnf1α and Ptf1a expression is associated with increased proliferation of cells involved in pancreatic repair/regeneration
To assess the kinetics of cell proliferation in relationship to changes in transcription factor expression, bromodeoxyuridine (BrdU) incorporation was quantified. Acinar cell proliferation increased at day 2, peaked at day 4, and returned to baseline levels by day 20 (figure 4). Of note, acinar BrdU labelling coincided with low expression of Hnf1α and Ptf1a (figure 2). Islet and ductal cell proliferation was negligible. By contrast, a marked increase in the proliferation of interstitial cells was detected as soon as 7 h after initiation of pancreatitis, reaching a peak at 24 h (when most proliferating cells were found in the interstitium or periacinar spaces), and returning to baseline at day 7 (figure 4). Double labelling for Ki67 and cell-specific markers showed that, at 24 h, 43.7% and 4.8% of Ki67+ cells expressed myeloperoxidase or desmin, respectively. Only a minor proportion of Ki67+ cells were lymphocytes (online supplementary figures 4 and 5).

Cell proliferation kinetics after pancreatitis induction. Bromodeoxyuridine (BrdU) incorporation at 24 h (A) or 4 days (B). At 24 h almost all proliferating cells are located in the interstitium or periacinar spaces. Four days after pancreatitis, the majority of proliferating cells are acinar. (C) Time-dependent plot of proliferating cells per high power field (×200) according to their location. There is a remarkable interstitial-to-acinar shift of proliferating cells over time.
Hnf1α- and Ptf1a-deficient mice show higher acinar cell proliferation rates than wild-type animals
The findings described above raised the possibility that cell proliferation changes might, in part, result from downregulation of Hnf1α and/or Ptf1a. In agreement with this notion, baseline acinar cell proliferation was higher in Hnf1α−/− and Ptf1a+/− mice (figure 5A) and was significantly higher in Hnf1α−/− mice—but not in Ptf1a+/− mice—during the regenerative phase of acute pancreatitis (figure 5A). Ptf1a has been shown to repress cell proliferation.28 Interestingly, Hnf1α−/− mice had reduced levels of Ptf1a protein, both at basal conditions and after pancreatitis (figures 5B,C), possibly contributing to the increased acinar proliferation rates.

Acinar cell proliferation and Ptf1a expression in wild-type and Hnf1α- and Ptf1a-deficient mice. (A) Number of pHH3-positive acinar cells per high power field (×200) in Hnf1α−/− mice, Ptf1a+/− mice and wild-type littermates. (B) Ptf1a expression in the pancreas of wild-type (+/+) and Hnf1α−/− mice. (C) Reduced immunostaining for Ptf1a protein in Hnf1α−/− mice.
Hnf1α−/− mice display altered Ptf1a expression and exocrine differentiation and respond to caerulein
To explore the notion that Hnf1α plays a role in acinar cell physiology, we examined the exocrine compartment of Hnf1α−/− mice. Hnf1α−/− mice were smaller than wild-type littermates (20.8±1.4 vs 31.6±2.5 g; p=0.002) and had smaller pancreata (229±15 vs 378±27 mg; p=0.0003), although the pancreas/body weight ratio remained constant (11.06±0.5 vs 12.01±0.4 mg/g; p=0.15) (n=8). At the histological level, the pancreas architecture of Hnf1α−/− mice was grossly normal. However, loose interacinar spaces, thin basophilic rim, and small peripheral nuclei were apparent in most acinar cells (figure 6A). Ultrastructural analysis revealed well-preserved acinar cells with somewhat dilated rough endoplasmic reticulum (figure 6A) and zymogen granules of normal size in the proximity of apical lumina. Interestingly, cells containing endocrine granules were identified inside acini from Hnf1α−/− mice, but not in wild-type littermates. In addition, both zymogen and endocrine granules were detected in isolated acinar cells, indicating that these cells display abnormal differentiation (figure 6A).

Pancreas histology, ultrastructure and function in Hnf1α−/− mice. (A) (i,ii) Loose interacinar spaces, thin basophilic rim, and small peripheral nuclei are observed in acinar cells of Hnf1α−/− mice (H&E; ×200). (iii,iv) Representative acinar cell electron micrographs from Hnf1α−/− mice (original magnification: 3400 for (iii) and 13 500 for (iv)). Arrows: dilated rough endoplasmic reticulum. Arrowheads: endocrine-like granules in an acinar cell. (B) Quantitative reverse transcriptase-PCR analysis of exocrine gene expression in Hnf1α−/− pancreata. (C) Reduced amylase activity in Hnf1α−/− pancreata. (D) Effects of caerulein on amylase secretion of isolated pancreatic acini from wild-type and Hnf1α−/− pancreata (n=5).
mRNA levels of most digestive enzymes were significantly lower in the pancreas of Hnf1α−/− mice (figure 6B), as were levels of Ptf1a, Mist1 and Rbpj transcripts. By contrast, Keratin 7, Pdx1 and Rbpjl mRNA levels were unaffected, supporting the selectivity of these changes.
Pancreatic amylase was reduced by 35% (p=0.019) (figure 6C) in homogenates from Hnf1α−/− pancreata. To assess the secretory response, isolated acini were challenged with caerulein in vitro (figure 6D): a biphasic dose-dependent amylase secretion response was observed in both wild-type and Hnf1α−/− mice. However, Hnf1α−/− mice exhibited a suboptimal response.
On induction of acute caerulein pancreatitis, Hnf1α−/− mice showed acute damage (online supplementary figure 6), which persisted focally by day 7 and was more pronounced after two bouts of pancreatitis. Induction of 3-week pancreatitis, to favour fibrosis development, resulted in high mortality in Hnf1α−/− mice.
Hnf1α−/− controls the expression of Nr5a2, an orphan nuclear receptor that regulates digestive enzyme gene expression
The exocrine defects observed in Hnf1α−/− mice suggested that digestive enzyme genes may be direct targets of Hnf1α. However, the search for putative binding sites using extensive information gathered from the in silico and genome-wide ChIP analysis of Hnf1α targets provided no evidence suggestive of Hnf1α being a direct regulator of digestive enzymes, Ptf1a, Rbpjl or Mist1.14 ,15 However, a high score consensus Hnf1 motif was identified at positions −237 to −222 in the Nr5a2 promoter that is also found in the promoter of the human gene. Nr5a2 is an orphan nuclear receptor, and there is evidence that it can regulate expression of some acinar genes.21 We assessed Nr5a2 mRNA levels in wild-type mice after caerulein pancreatitis and in Hnf1α−/− mice (figures 7A,B), two experimental conditions associated with reduced acinar gene expression and function. In both situations Nr5a2 transcripts were markedly reduced. Indeed, during caerulein pancreatitis, Nr5a2 mRNA levels followed a pattern similar to that of digestive enzyme mRNAs (figure 7A). To determine whether Hnf1α can regulate Nr5a2, promoter reporter and ChIP assays were performed. Transfection of the Hnf1α cDNA led to an increased activity of an Nr5a2 promoter reporter construct, and mutation of the putative Hnf1α-binding site abolished this effect (figure 7C). Using ChIP, a clear enrichment of the sequence containing the evolutionarily conserved Hnf1 motif, but not of an unrelated intronic sequence, was present in the immunoprecipitates detected using antibodies that recognise Hnf1α (figure 7D).
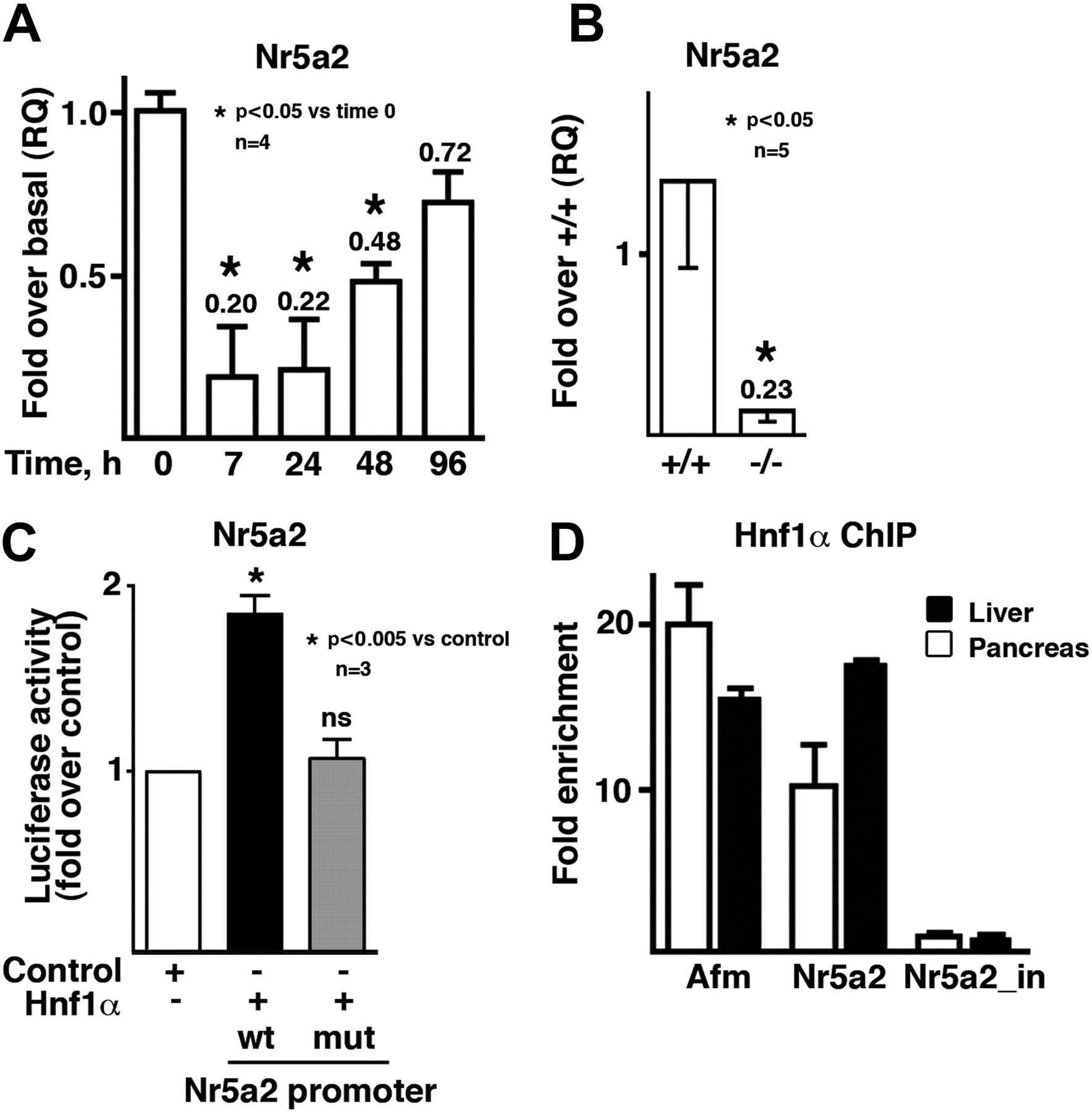
Nr5a2 is a direct target of Hnf1α in the pancreas and its expression is altered in Hnf1α−/− mice. (A) Reduced pancreatic Nr5a2 expression during caerulein pancreatitis. (B) Reduced Nr5a2 transcript expression in the pancreas of Hnf1α−/− mice. (C) Nr5a2 promoter activity is regulated by Hnf1α and depends on a putative Hnf1α motif at positions −237 to −222. Nr5a2-luciferase reporter activity of the −1.2 kb promoter fragment (using both wild-type and mutant forms for Hnf1α-binding site) in HEK293 cells. Ns, not significant. (D) Hnf1α binds to the promoter of Nr5a2 and a control target gene (afamine, Afm) but not to an Nr5a2 intronic sequence.
Discussion
There is increasing interest in dissecting the transcriptional circuits that maintain differentiated cell phenotypes in the adult pancreas. Knowledge of the transcriptional control of pancreatic cell dynamics in response to injury is key to understanding the events involved in pancreatic regeneration, the development of chronic pancreatitis, and pancreatic carcinogenesis. Here, we provide evidence for injury-responsive regulated expression of Ptf1a, Pdx1 and Hnf1α in acinar and endocrine cells. We also provide evidence for transient deranged expression of genes under the control of these transcription factors, resulting in inappropriate pancreatic function.
Caerulein-induced acute pancreatitis results in histological changes affecting acinar cells, but no gross abnormality is present in islets. The extent and duration of reduced enzyme gene expression cannot be merely explained by the cell destruction derived from pancreatitis, since morphological examination of pancreatic specimens reveals neither profound nor prolonged acinar cell necrosis.
We have previously reported a marked downregulation of Ptf1a in rats after caerulein pancreatitis,26 preceding a sharp reduction in pancreatic amylase content and activity. In that model, Ptf1a is restored during the initial phase of pancreatic regeneration, before the complete recovery in pancreatic amylase content occurs. Ciclosporin delays the recovery of nuclear Ptf1a expression, probably preventing regeneration and promoting development of chronic-like pancreatitis.26 On caerulein pancreatitis, the Notch pathway, which represses PTF1 function and acinar differentiation during development,29 is strongly activated.19 ,20 Ptf1a is transiently reduced, followed by a reduction in expression of PTF1 target genes such as amylase and elastase. Ptf1a recovery is not immediately followed by normalisation of digestive enzyme mRNA levels, possibly because of the requirement of additional transcription factors.
Several reports indicate that pancreatic exocrine function is impaired after acute pancreatitis in humans and that it tends to recover at later stages.30–33 Our finding that expression of exocrine transcription factors and enzymes is reduced after mild pancreatitis adds new insights that may help to explain the exocrine function impairment.
Despite the lack of morphological changes in islets, endocrine genes are also downregulated during caerulein pancreatitis, as are endocrine transcription factors, in association with a transient reduction in plasma insulin and an abnormal response to oral glucose overload. This effect is in agreement with clinical observations of reduced exocrine and endocrine responses in humans after acute pancreatitis.30–32 ,34 Patients with acute pancreatitis may present with hyperglycaemia (included in the Ranson score for prediction of severity), which usually subsides as pancreatitis resolves. Acute pancreatitis-associated hyperglycaemia has been classically attributed to insulin resistance or to loss of pancreatic tissue.33 ,35
The occurrence of both exocrine and endocrine dysfunction in acute caerulein-induced pancreatitis, together with downregulation of Hnf1α, led us to explore whether Hnf1α may play a role in acinar cells. The relevance of Hnf1α in the endocrine pancreas is underlined by the fact that Hnf1α mutations are responsible for MODY3, a disease characterised by compromised insulin secretion. In mice, combined deficiency of both Hnf1α and Pdx1 results in increased reduction in insulin expression and greater glucose intolerance than the isolated deficiency of either transcription factor.36 We find that, in response to pancreatitis, a combined transient decrease in Hnf1α and Pdx1 expression ensues. We also show reduced insulin expression (mRNA and protein) and consequent transient glucose intolerance. Insulin pre-mRNA accurately reflects immediate changes in transcriptional rates.37 The reduction in insulin pre-mRNA in a time frame when Pdx1 and Hnf1α are markedly diminished is of similar magnitude to the reduction in insulin mRNA reported by Shih et al in mice with combined deficiency of Hnf1α and Pdx1 (46%).36 Because of its longer half-life, assessment of mature insulin mRNA may have hampered detection of expression changes. A broader effect on the endocrine compartment is supported by the reduced expression of somatostatin, pancreatic polypeptide and glucagon. Caerulein can promote the development of pancreatic cancer driven by mutant K-ras in mice,38 ,39 but the mechanisms involved have not been completely unravelled. Our findings suggest that the effects of caerulein outside of the acinar compartment may also contribute to tumour promotion, especially in the context of K-ras activation in β cells.39 Importantly, we also show that different models of acute experimental pancreatitis result in distinct effects on the endocrine compartment.
The effects of caerulein on cell differentiation are accompanied by changes in proliferation, as previously reported: Ptf1a has antiproliferative effects in vitro and acinar cells from Ptf1a+/− mice display increased proliferation.28 We find increased proliferation coinciding with reduced nuclear Ptf1a and Hnf1α in acinar cells, in agreement with previous work and with reports on Notch activation after acute pancreatitis.19 ,20 ,40 We also confirm increased acinar proliferation in Ptf1a+/− and Hnf1α−/− mice in basal conditions and—in the latter—after acute pancreatitis. Hnf1α suppresses cell proliferation in hepatocytes, as in acinar cells, and it enhances proliferation in islets.15 The role of Hnf1α as a regulator of acinar cell proliferation merits further study.
The observations described above prompted us to examine the acinar transcriptome in Hnf1α−/− mice. qRT-PCR analyses indicate a reduced expression of transcripts coding for digestive enzymes, Ptf1a and Mist1. In addition, Hnf1α−/− mice exhibit reduced pancreatic amylase activity, subtle ultrastructural changes in acinar cells including the concomitant occurrence of exocrine and endocrine granules, and a suboptimal acinar secretory response in vitro. Defective specification and increased crypt proliferation have been reported in the intestine of Hnf1α−/− mice.41
To decipher the mechanisms involved in defective exocrine function in Hnf1α−/− mice, we analysed the promoters of deregulated genes and used published global Hnf1α occupancy studies. We found no evidence for a direct regulation of acinar transcription factors or digestive enzymes by Hnf1α. Bioinformatic analyses and published genome-wide ChIP data pointed to Nr5a2 as a candidate Hnf1α direct target in the pancreas, which was confirmed by promoter reporter and ChIP/qPCR assays.14 ,15 Accordingly, Nr5a2 mRNA levels are markedly reduced in Hnf1α−/− and wild-type mice after caerulein pancreatitis. These effects are selective for acinar cells, since Nr5a2 expression is not dependent on Hnf1α in the liver despite being a direct target also in this tissue, and Nr5a2 transcripts and protein are low/undetectable in islets (not shown).15 Furthermore, Hnf1α expression in isolated stellate cells is low/undetectable by both immunocytochemistry and RT-qPCR (not shown).
Hnf1α has previously been implicated in the regulation of many genes in hepatocytes and islets, where it is thought to play a role in maintaining cell differentiation.15 However, this is the first report to assign regulatory functions to Hnf1α in acinar physiology. Our data suggest that Hnf1α promotes acinar differentiation through the regulation of Nr5a2, which codes for an orphan nuclear receptor expressed in endoderm-derived tissues that is required for normal embryonic development.42 Nr5a2 is directly regulated by Pdx1 during pancreatic development and it is expressed at the tip of pancreatic branches at e14.5.43 ,44 In turn, Nr5a2 is a direct regulator of carboxyl ester lipase,22 and recent work indicates a more general role for this protein in the regulation of pancreatic exocrine function.45 While we show here that Hnf1α regulates Nr5a2, there is evidence that both proteins can cooperate in the regulation of target genes in the liver and that they interact physically,46 ,47 providing additional mechanisms of regulatory interaction. A working model for the participation of Hnf1α in the acinar transcriptional regulatory network is shown in figure 8.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hnf1α and the acinar transcription factor regulatory network: a working model. As shown in this work, Hnf1α regulates Nr5a2 expression. Rbpjl, a PTF1 complex component that is required for full maturation of acinar cells, is regulated by both Nr5a2 and Ptf1a.45 PTF1 also regulates Pdx1, which, in turn, regulates Nr5a2. Regulatory interactions established in non-pancreatic cells are shown as dashed lines. Transcription factors binding the promoter region of acinar genes are shown encircled in the bottom panel.
Overall, we identify novel components of the acinar transcription factor network. We find that pancreatic exocrine and endocrine functions are closely inter-related during caerulein-induced pancreatitis: Hnf1α can be added to the list of transcription factors involved in both pancreatic components. A complex network of regulatory interactions emerges, since Hnf1α regulates Pdx1 expression which—in turn—can regulate Ptf1a and the latter can also regulate Pdx1.7 ,48 ,49 Our findings also support the need of a more detailed analysis of exocrine function in patients with MODY3 (particularly during the recovery phase of acute injury) and, more generally, in endocrine disorders. Our observations may be relevant to understanding how chronic pancreatitis and diabetes associate with an increased risk of pancreatic cancer. Interestingly, genetic variation in Nr5a2 and Hnf1α loci has recently been implicated in susceptibility to developing pancreatic ductal adenocarcinoma.50 ,51
Acknowledgments
We thank J Pujal, A Pinho, E Sampere and N del Pozo for technical assistance, M Pontoglio for providing Hnf1α antibodies, K Schoonjans for providing Nr5a2 promoter reporter constructs, J Lloreta for help with the ultrastructural analyses, and the investigators mentioned in the text for reagents or mice.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
Footnotes
Funding This work was supported, in part, by: grants PI080342 from Fondo de Investigación Sanitaria and Acción Especial de Genómica (GEN2001-4748-c05-03) to XM; grant PI080280 from Fondo de Investigación Sanitaria to ECV; grants SAF2004-01137, SAF2007-60860, Acción Especial de Genómica (GEN2001-4748-c05-01), and ONCOBIO Consolider from Ministerio de Ciencia e Innovación (Madrid, Spain) to FXR; grant PI080511 from Fondo de Investigación Sanitaria and Acción Especial de Genómica (GEN2001-4748-C05-04) to AS; US Public Health Service grant DK061220 to RJM; and Acción Especial de Genómica (GEN2001-4748-c05-02) to JF. AM was supported by a Juan de la Cierva contract from Ministerio de Ciencia y Tecnología.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement We will provide any data that is cited in the text upon request to XM or FXR.