Article Text

Abstract
Objective Oesophagitis might result from the effects of chemokines produced by oesophageal cells in response to gastro-oesophageal reflux, and not solely from the direct, caustic effects of refluxed gastric juice. Proton pump inhibitors (PPI) can block chemokine production through mechanisms independent of their antisecretory effects. We studied omeprazole effects on chemokine production by oesophageal epithelial cells exposed to acidic bile salts.
Design Human primary and telomerase-immortalised oesophageal squamous cells were exposed to acidic bile salt medium with or without omeprazole pretreatment. Interleukin (IL)-8 expression was determined by RT-PCR and ELISA. IL-8 promoter activity was measured by luciferase reporter assay. Binding of NF-κB and AP-1 subunits to the IL-8 promoter was assessed by chromatin immunoprecipitation (ChIP) assay. Immune cell migration induced by conditioned medium was determined by a double-chamber migration assay system.
Results Acidic bile salt medium caused oesophageal epithelial cells to express IL-8 mRNA and protein by activating the IL-8 promoter through NF-κB and AP-1 binding. Omeprazole inhibited that acidic bile salt-stimulated IL-8 expression by blocking the nuclear translocation of p65 (an NF-κB subunit), and by blocking the binding of p65, c-jun and c-fos (AP-1 subunits) to the IL-8 promoter. Omeprazole also blocked the ability of conditioned medium from cells exposed to acidic bile salts to induce immune cell migration.
Conclusions In oesophageal squamous epithelial cells, omeprazole inhibits IL-8 expression through effects on NF-κB and AP-1 that are entirely independent of effects on gastric acid secretion. These previously unrecognised PPI effects might contribute to the healing of reflux oesophagitis.
- GASTROESOPHAGEAL REFLUX DISEASE
- PROTON PUMP INHIBITION
- CHEMOKINES
- EPITHELIAL CELLS
- MOLECULAR MECHANISMS
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
-
It has been widely assumed that proton pump inhibitors (PPI) heal gastro-oesophageal reflux disease (GORD) solely because they inhibit gastric acid secretion.
-
Recent studies suggest that reflux oesophagitis might not result solely from the direct, caustic effects of refluxed acid on oesophageal epithelial cells, but rather from the effects of chemokines such as interleukin (IL)-8 produced by those cells in response to the gastro-oesophageal reflux of acid and bile.
-
PPIs have a number of potential anti-inflammatory actions that are independent of their effects on gastric acid secretion, including the ability to inhibit chemokine production in certain epithelial cells.
What are the new findings?
-
Cultures of primary and telomerase-immortalised, non-neoplastic oesophageal squamous cells exposed to acidic bile salts secrete IL-8, which is a major mediator of inflammation.
-
Omeprazole blocks the acidic bile salt-stimulated IL-8 secretion in oesophageal squamous cells by blocking nuclear translocation of p65 (an NF-κB subunit) and by decreasing binding of p65, c-jun and c-fos (AP-1 subunits) to the IL-8 promoter.
-
Omeprazole pretreatment blocks the ability of conditioned medium from oesophageal squamous cells exposed to acidic bile salts to induce immune cell migration.
How might it impact on clinical practice in the foreseeable future?
-
We have demonstrated anti-inflammatory effects of PPIs that are entirely independent of their effects on gastric acid secretion, and that might contribute to the beneficial effects of PPIs in the treatment of GORD.
-
These findings suggest a potential role for chemokine-directed therapies in the treatment of GORD.
Introduction
Proton pump inhibitors (PPIs are the most effective medications available for the treatment of gastro-oesophageal reflux disease (GORD), and PPIs are considered the medical therapy of choice for erosive reflux oesophagitis.1 The PPIs are so named because they inhibit H+, K+ATPase, the ‘proton pump’ of the parietal cell that is responsible for gastric acid secretion. It has been assumed widely that gastric acid inhibition is the only important beneficial action of the PPIs. According to this prevailing notion, PPIs heal reflux oesophagitis solely because they decrease the production of gastric acid, which reduces acid reflux, which prevents acid–peptic damage to the oesophagus. However, PPIs have been found to have a number of anti-inflammatory actions that are independent of their effects on gastric acid secretion.2 In human gastric cancer cells and human umbilical vein epithelial (HUVEC) cells, for example, PPIs inhibit the production of chemokines including interleukin (IL)-8, which is a major mediator of inflammation.3 PPIs also decrease adhesion molecule production by endothelial and inflammatory cells4–7 and, in oesophageal squamous cells from patients with eosinophilic oesophagitis, we have shown that PPIs inhibit the production of eotaxin-3 (a chemokine that attracts eosinophils) stimulated by exposure to Th2 cytokines.8 ,9 Conceivably, such anti-inflammatory effects of PPIs might contribute to their beneficial actions in GORD.
Another traditional notion about GORD that might be incorrect is that reflux oesophagitis is the direct result of a caustic, chemical injury inflicted on the oesophagus by refluxed gastric acid. According to this traditional notion, reflux oesophagitis begins with acid and pepsin attacking and destroying squamous cells at the oesophageal luminal surface, progressing later to deeper mucosal layers as acid burns through the lining of the oesophagus.10 Our recent studies using a rat model of reflux oesophagitis have challenged this concept of GORD pathogenesis, however.11 After the surgical induction of reflux by oesophagoduodenostomy in the rat, we have shown that reflux oesophagitis starts with a lymphocytic infiltration in the oesophageal submucosa, which progresses over the course of weeks to involve the mucosal surface. We also have found that human oesophageal squamous cells exposed to an acidic bile salt medium (similar in composition to gastric juice) secrete chemokines, including IL-8, which can induce the migration of inflammatory cells.11 Based on these findings, we have proposed a new hypothesis for the development of reflux oesophagitis in which the reflux of gastric juice stimulates oesophageal squamous cells to secrete chemokines that attract inflammatory cells. According to this hypothesis, it is chemokine-mediated inflammation, not an acid burn, that initially damages the oesophageal mucosa in patients with GORD.
Noting that oesophagitis might be mediated by chemokines (like IL-8) produced by the oesophagus in response to gastro-oesophageal reflux, and that PPIs can block chemokine production through acid-independent mechanisms, we hypothesised that PPIs might interfere with reflux-stimulated chemokine production by oesophageal epithelial cells, and that this acid-independent mechanism might contribute to the efficacy of PPI therapy in GORD. To explore this hypothesis, we studied the effects of omeprazole on IL-8 secretion induced by exposure to acidic bile salts in primary oesophageal squamous epithelial cells and in telomerase-immortalised, non-neoplastic oesophageal squamous cell lines.
Materials and methods
Patients providing oesophageal biopsy specimens for primary cell cultures
These studies were approved by the Institutional Review Board on Human Studies of the Dallas VA Medical Center. Study subjects were patients scheduled for elective endoscopy who provided written, informed consent for participation. Six patients with GORD (5 men, average age 56.4±4.6 years) and three control subjects (all men, average age 54.3±5.4 years) who had no symptoms or endoscopic signs of oesophageal disease had biopsies taken from the distal, squamous-lined oesophagus to establish primary cell cultures as previously described.8 Briefly, primary cell cultures are established by placing oesophageal biopsy specimens essentially unaltered into culture medium. The diagnosis of GORD was based on one or any combination of the following: (1) heartburn (defined as an uncomfortable, retrosternal burning sensation relieved by acid suppression) for at least 3 months, (2) the endoscopic finding of erosive oesophagitis (Los Angeles grades A–C) and (3) long-segment Barrett's oesophagus (defined as >3 cm of biopsy-confirmed oesophageal-specialised intestinal metaplasia).
Cell lines
We used non-neoplastic, telomerase-immortalised oesophageal squamous (NES) cell lines created from endoscopic biopsy specimens of the distal oesophagus from two patients who had GORD with Barrett's oesophagus (NES-B3T, NES-B10T) and from two who had GORD without Barrett's oesophagus (NES-G2T, NES-G4T).12 ,13 All experiments involved only oesophageal squamous cells, not Barrett's columnar cells. We also used immune cell lines, which included a human T-cell line (Jurkat) and a human neutrophil line (HL-60 promyelocytic leukaemia cells differentiated with dimethyl sulfoxide) (American Type Culture Collection, Manassas, Virginia, USA) as previously described (see online supplementary methods).11 ,14 ,15
Acidic bile salt exposure and omeprazole treatment
For individual experiments, NES cell lines and primary oesophageal squamous cells were cultured either in neutral medium (pH 7.2) alone or in neutral medium with periodic exposures to acidic medium (pH 4.0) containing a mixture of conjugated bile acids (glycocholic acid, glycodeoxycholic acid (both from Calbiochem, San Diego, California, USA), taurocholic acid, glycochenodeoxycholic acid, taurochenodeoxycholic acid and taurodeoxycholic acid (Sigma–Aldrich, St Louis, Missouri, USA) in a 20:6:3:15:3:1 molar concentration, total concentration 400 μM) designed to simulate gastric refluxate found in patients with GORD and Barrett's oesophagus.16 ,17 Neutral or acidic bile salt medium was added for 10 min, three times a day, for a total of 5 days unless otherwise indicated. These exposure durations were chosen to simulate typical episodes of gastro-oesophageal reflux.18 We used omeprazole at a concentration of 35 μM (gift of AstraZenca R&D, Molndal, Sweden, and also purchased commercially from Sigma,8 ,9 St Louis, Missouri, USA), which was added to the medium overnight after the third and fourth days of acidic bile salt treatment.
For experiments using chromatin immunoprecipitation (ChIP) and IL-8 promoter activity assays to evaluate subunit binding to the IL-8 promoter, cell lines were treated with acidic bile salt or control medium for 24 h, with or without 24 h of pretreatment by omeprazole in neutral pH medium. After pretreatment, the neutral medium was removed and replaced with either control medium (with omeprazole) or acidic bile salt medium (pH 5.5 with omeprazole) for the duration of the experiment.
Semiquantitative reverse transcription PCR
Semiquantitative PCR was performed for IL-8 and GAPDH mRNAs in NES cell lines and primary oesophageal squamous cells after day 5 of acidic bile salt exposure (see online supplementary methods).
ELISA for IL-8
We performed ELISA of conditioned medium from NES cells collected the morning after each day of exposure for 5 days, using commercially available ELISA kits (R&D Systems) to assess production of IL-8 (see online supplementary methods).
IL-8 promoter activity assays
For transfection studies, we used plasmid constructs containing the −162/+44 fragment of the human IL-8 promoter cloned into pGL3 upstream of a luciferase reporter (−162/+44 hIL-8/Luc), and −162/+44 hIL-8/Luc containing point mutations in the NF-κB and AP-1 DNA binding sites (ΔNF-κB 162/+44 hIL-8/Luc and ΔAP-1 162/+44 hIL-8/Luc; all plasmids were the generous gift of Dr Marc Hershenson, University of Michigan, Ann Arbor, Michigan, USA); pGL-3 Basic Luciferase Vector (Promega, Madison, Wisconsin, USA) lacking the IL-8 promoter insert served as a control.19 The renilla reporter pRL (Promega) plasmid was used to equalise for transfection efficiency. Data were expressed as relative light units for firefly luciferase normalised to renilla luciferase (see online supplementary methods).
Nuclear and cytoplasmic fractionation and western blot
For p65, c-jun and c-fos, NES cell lines were exposed to acidic bile salt or control medium for 2 h with or without omeprazole pretreatment for 24 h. The medium was removed and the cells were washed in cold phosphate-buffered saline. Nuclear extracts were isolated from cell lines using the NE-PER Nuclear and Cytoplasmic Extraction kit (Thermo Fisher Scientific, Rockford, Illinois, USA) per the manufacturer's instructions. Proteins were separated by SDS- polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes, and incubated with primary antibodies to total and phospho-p65, c-jun, c-fos, tubulin and TF-IID (see online supplementary methods).
ChIP assay
NES cell lines were treated with acidic bile salt or control medium for 24 h with or without omeprazole pretreatment, and a ChIP assay was performed as previously described (see online supplementary methods).12
Immune cell migration
We used a transwell system in which immune cells loaded into one chamber were observed to migrate into another. For each cell line, 5×105 cells were placed in the upper chamber of 24-well dual chamber culture plates separated by a polycarbonate-coated membrane with 5.0 μm pores (Corning, Lowell, Massachusetts, USA). The immune cell lines were serum starved overnight. Conditioned media from NES-G4T and NES-B10T cells treated with 4 or 5 days of neutral or acidic bile salt medium with and without omeprazole were added to the lower chamber as a chemotactic reagent. Immune cells were loaded into the upper chamber. After a 3 h incubation at 37°C, the upper chamber was removed and cells in the lower chamber were collected and counted.
Data analyses
Quantitative data are expressed as the mean±SEM. Statistical analyses were performed using a paired or unpaired Student t test with the Instat for Windows statistical software package (GraphPad Software, San Diego, California, USA). For multiple comparisons, an analysis of variance and the Student–Newman–Keuls multiple-comparisons test was performed using the Instat for Windows statistical software package (GraphPad). p Values ≤0.05 were considered significant for all analyses.
Results
Acidic bile salts induce IL-8 secretion in oesophageal squamous cell lines from GORD patients with and without Barrett's oesophagus
In earlier studies, we had demonstrated that acidic bile salts increased IL-8 secretion in NES-B3T and NES-B10T oesophageal squamous cell lines, which are derived from patients with Barrett's oesophagus.11 Now, we exposed the NES-G2T and NES-G4 squamous cell lines (derived from patients who had GORD without Barrett's oesophagus) to acidic bile salts, and determined IL-8 secretion over 5 days. As in the NES-B3T and NES-B10T cell lines,11 we found minimal baseline secretion of IL-8 in NES-G2T and NES-G4T (figure 1). Also like the squamous cells from Barrett's patients, acidic bile salt exposure significantly increased IL-8 secretion after 1 and 2 days in NES-G2T and NES-G4T, respectively, and IL-8 secretion continued to increase through day 5 (figure 1). Thus, acidic bile salts induce IL-8 secretion in oesophageal squamous cell lines from GORD patients, irrespective of whether they have Barrett's oesophagus.
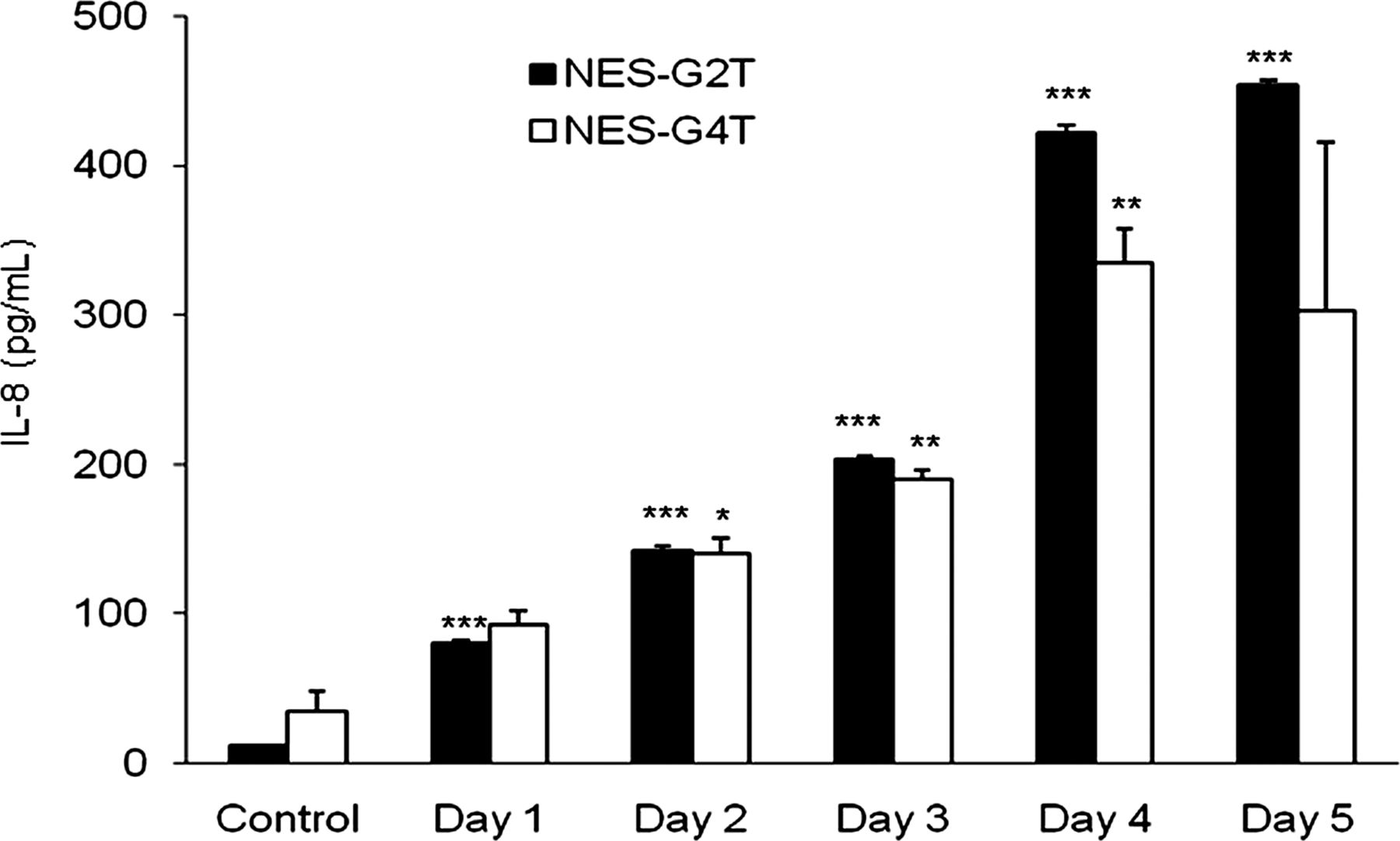
Oesophageal squamous cell lines from patients who have gastro-oesophageal reflux disease without Barrett's oesophagus (NES-G2T and NES-G4T) increase IL-8 protein secretion following exposure to acidic bile salt medium. The bar graphs depict the mean+SEM of at least 2 individual experiments. (*, p<0.05 compared with corresponding control; **, p<0.01 compared with corresponding control; ***, p<0.001 compared with corresponding control).
Acidic bile salts induce IL-8 secretion in primary oesophageal squamous cells from GORD patients
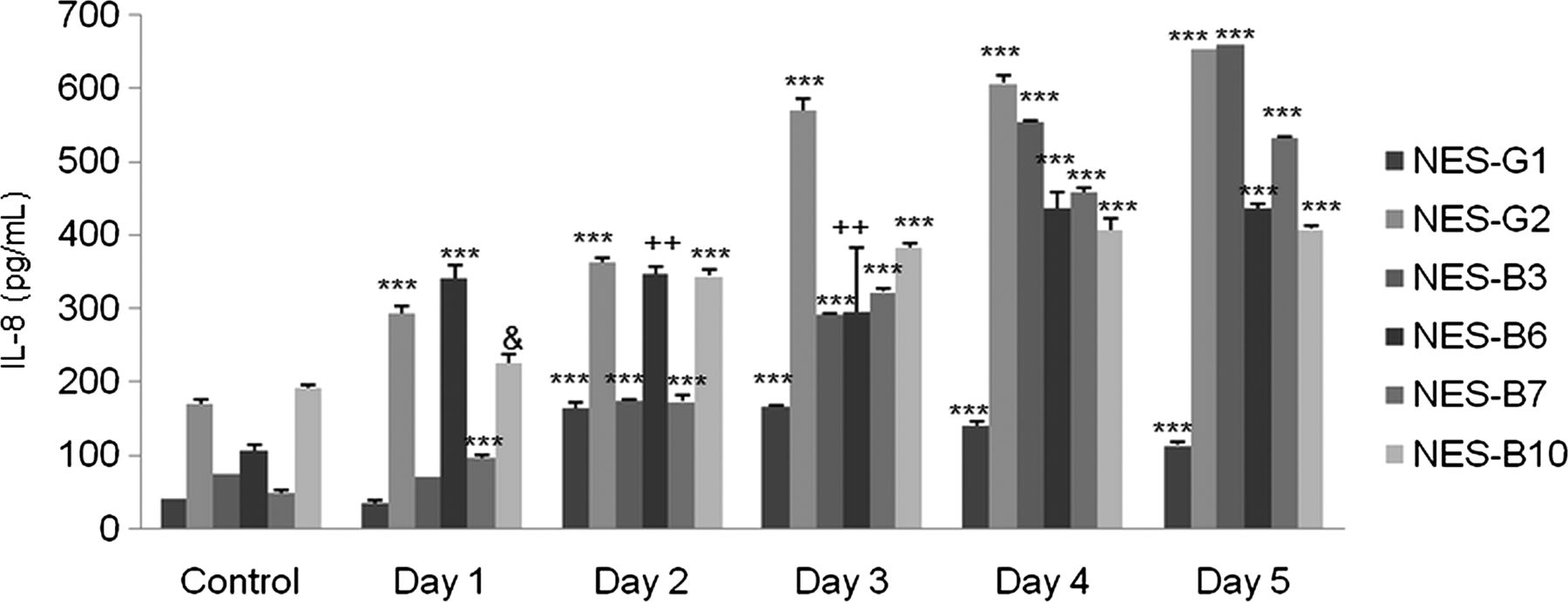
We next studied IL-8 secretion in response to acidic bile salt exposure in primary oesophageal squamous cells from two patients who had GORD without Barrett's oesophagus (NES-G1, NES-G2), and from four patients who had GORD with Barrett's oesophagus (NES-B3, NES-B6, NES-B7, NES-B10). We found that acidic bile salt exposure significantly increased IL-8 secretion in all the primary cells up to day 5, with wide variation among cells in IL-8 secretion levels (figure 2). These finding suggest that acidic bile salts increase IL-8 secretion in oesophageal squamous cells from patients with GORD, with considerable variation among individuals in their levels of stimulated IL-8 secretion.
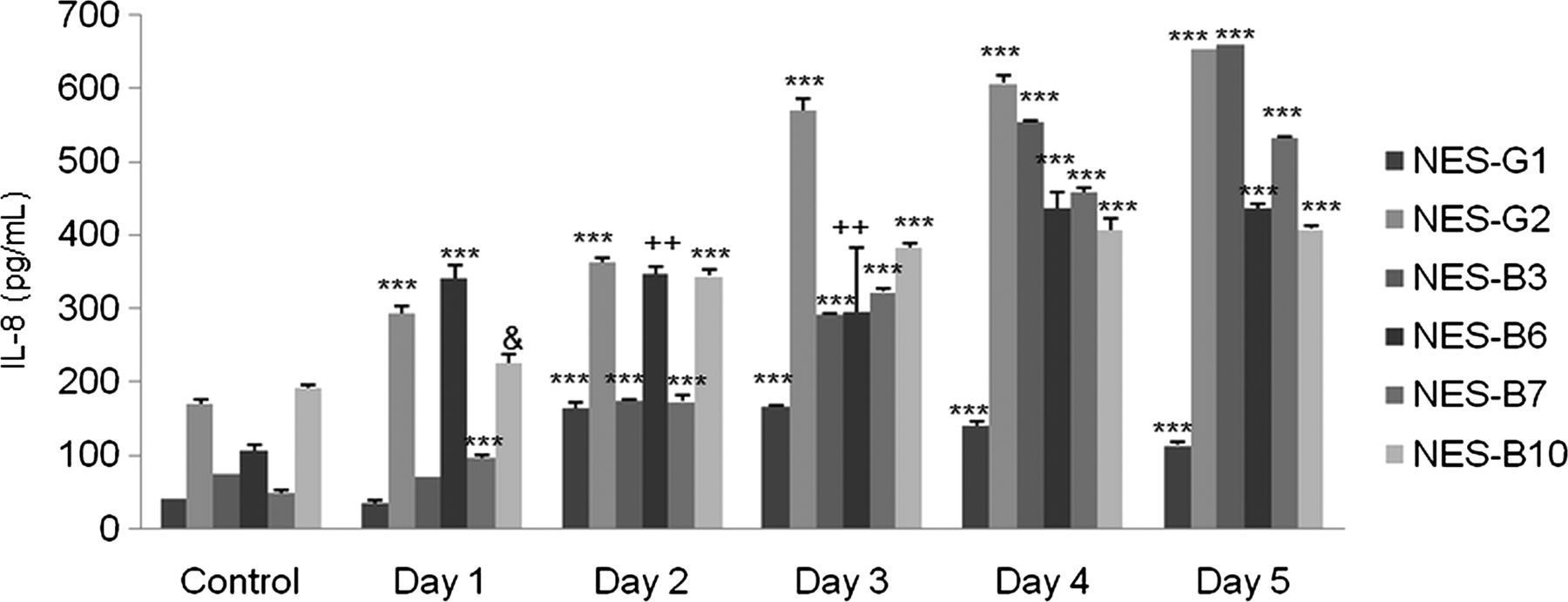
Primary oesophageal squamous (NES) cells from 6 gastro-oesophageal reflux disease patients all demonstrate increases in IL-8 protein secretion following exposure to acidic bile salt medium. The bar graphs depict the mean+SEM of at least 2 individual experiments. (&, p<0.05 compared with corresponding control; ++, p<0.01 compared with corresponding control; ***, p<0.001 compared with corresponding control).
Omeprazole blocks acidic bile salt-induced IL-8 secretion in oesophageal squamous cells and cell lines from GORD patients
We next explored effects of omeprazole on IL-8 protein secretion in oesophageal squamous cell lines and primary squamous cell cultures. In preliminary studies, we found that treatment with omeprazole alone had no apparent effect on the low levels of baseline secretion of IL-8 in oesophageal squamous cells (data not shown). We treated cells with acidic bile salt medium alone for 3 days, and then added omeprazole to the acidic bile salt medium for days 4 and 5. In all four NES cell lines, omeprazole significantly decreased IL-8 secretion in conditioned media on days 4 and 5 (figure 3A–D). In the primary oesophageal squamous cells, omeprazole caused a decrease in the mean level of IL-8 secretion on days 4 and 5, but the decrease achieved statistical significance only on day 4 (figure 3E,F).

Omeprazole (Ome) blocks IL-8 protein secretion induced by acidic bile salt medium in NES cell lines and primary NES cells. (A) NES-B10T, (B) NES-B3T, (C) NES-G4T, and (D) NES-G2T were exposed to acidic bile salt medium (A&B) for 4 or 5 days in the presence or absence of omeprazole . Data are the mean+SEM of 2 separate experiments. (***, p≤0.001 compared with corresponding control; ###, p≤0.001 compared with corresponding A&B stimulation alone) (E) Primary NES cells from 6 patient with gastro-oesophageal reflux disease exposed to A&B for 4 days or (F) for 5 days in the presence or absence of omeprazole. Horizontal line in each column represents the mean for all 6 patients.
Omeprazole blocks acidic bile salt-induced IL-8 secretion in oesophageal squamous cells from control subjects without oesophageal disease
To determine if PPI inhibition of acidic bile salt-stimulated cytokine production is a feature limited to the diseased oesophagus of GORD patients, we studied the effects of omeprazole on IL-8 protein secretion in primary oesophageal squamous cells from three control subjects without oesophageal disease (NES-N1, NES-N2, NES-N3). As in the GORD cells, exposure to acidic bile salts significantly increased IL-8 secretion at days 4 and 5 in all three control cells, and omeprazole significantly decreased that IL-8 secretion (figure 4).
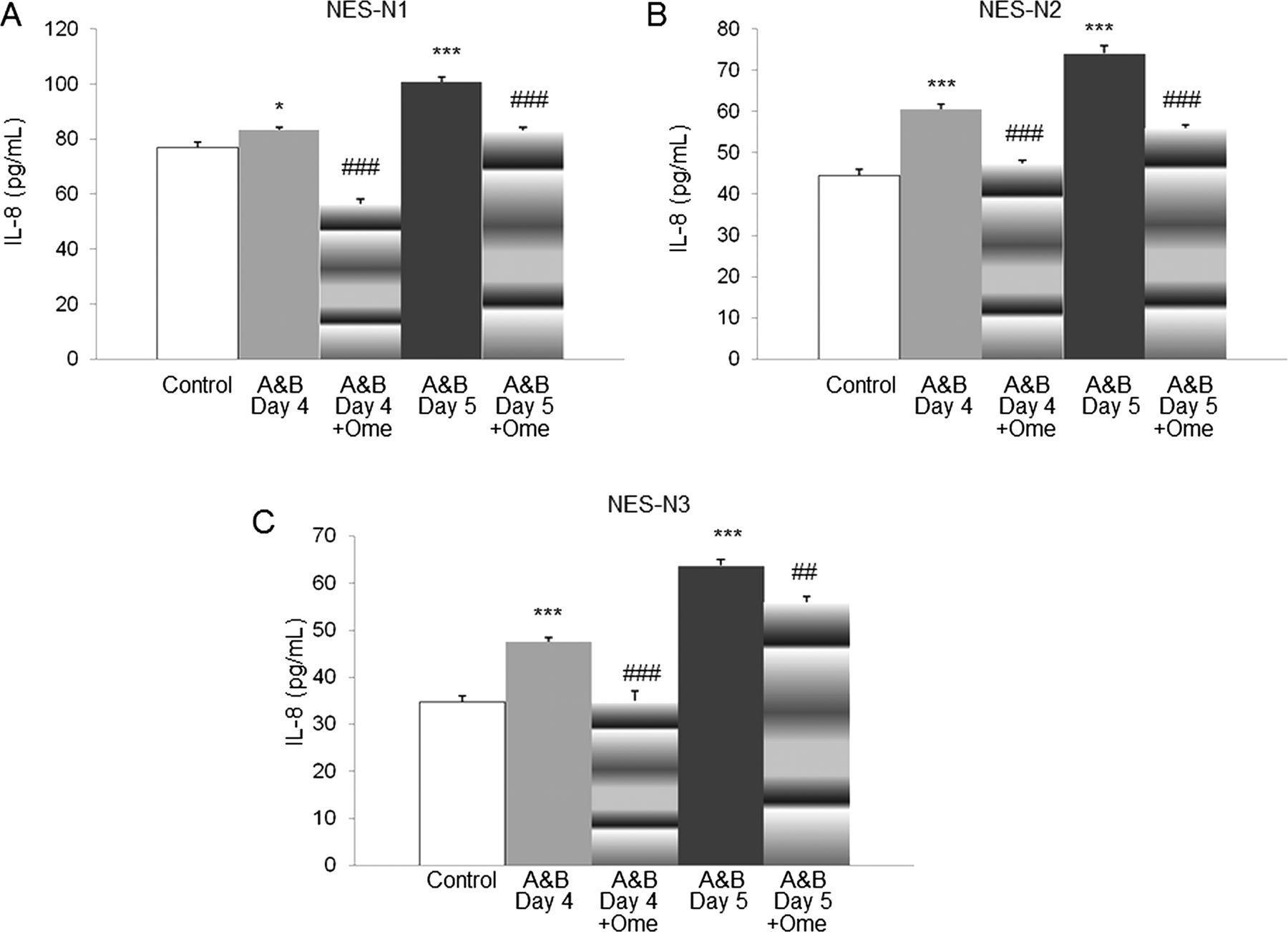
Omeprazole (Ome) blocks IL-8 protein secretion induced by acidic bile salt medium in primary oesophageal squamous cells from control subjects without oesophageal disease. (A) NES-N1, (B) NES-N2, and (C) NES-N3 were exposed to acidic bile salt medium (A&B) for 4 or 5 days in the presence or absence of omeprazole . Data are the mean+SEM of 2 separate experiments. (*, p<0.05 compared with corresponding control; ***, p≤0.001 compared with corresponding control; ##, p≤0.01 compared with corresponding A&B stimulation alone; ###, p≤0.001 compared with corresponding A&B stimulation alone).
Omeprazole suppresses acidic bile salt-stimulated IL-8 mRNA expression and promoter activation in oesophageal squamous cells and cell lines from GORD patients
We next explored the mechanisms underlying how omeprazole blocks acidic bile salt-stimulated IL-8 protein secretion. To determine whether omeprazole affected acidic bile salt-induced IL-8 transcriptional regulation, RT-PCR was performed to evaluate IL-8 mRNA expression in all four NES cell lines and in the five primary cell cultures that demonstrated decreased IL-8 protein secretion with omeprazole in acidic bile salt medium. Compared to cells stimulated with acidic bile salts alone, omeprazole caused a significant decrease in IL-8 mRNA levels for all NES cell lines (figure 5A) and primary cells (figure 5B).
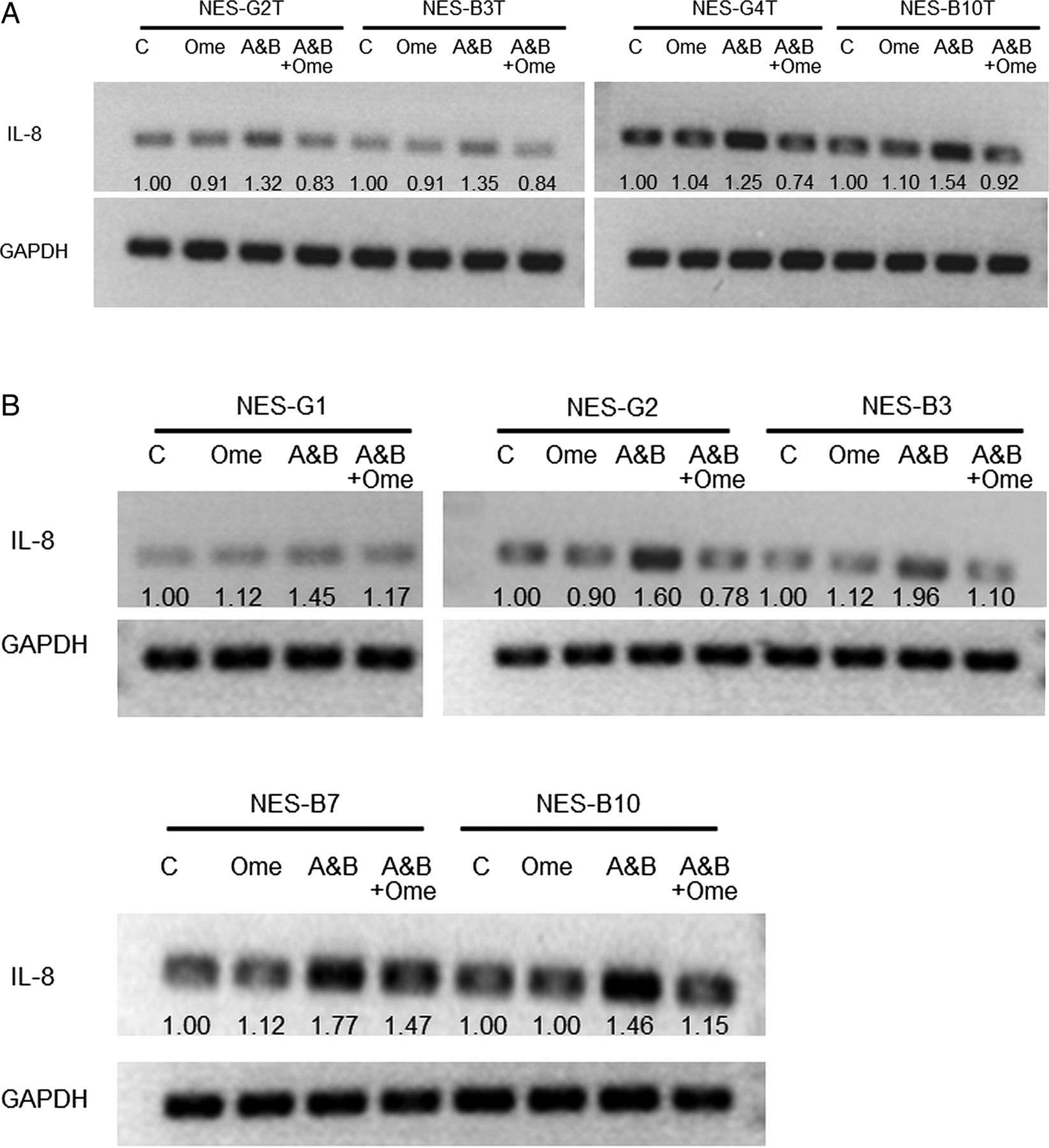
Omeprazole (Ome) decreases IL-8 mRNA expression induced by acidic bile salt medium in (A) NES cell lines and (B) primary NES cells. NES cell lines and primary NES cells were exposed to acidic bile salt media (A&B) for 5 days in the presence or absence of omeprazole (35 μM). Numbers represent normalised densitometric values. Depicted is 1 of 2 separate experiments.
Using cell lines, we next determined whether acidic bile salts activate the IL-8 promoter in NES cells. We transiently transfected the cells using the −162/+44 hIL-8/Luc promoter. After stimulation with acidic bile salts for 24 h, all four NES cells exhibited significantly increased IL-8 promoter activity (figure 6A–D). We next treated NES-B10T and NES-G4T cells with acidic bile salts in the presence of omeprazole. We found that omeprazole alone had no effect on IL-8 promoter activity. Compared to cells stimulated with acidic bile salts alone, we found a significant decrease in IL-8 promoter activity in NES cell lines treated with omeprazole (figure 6E,F).
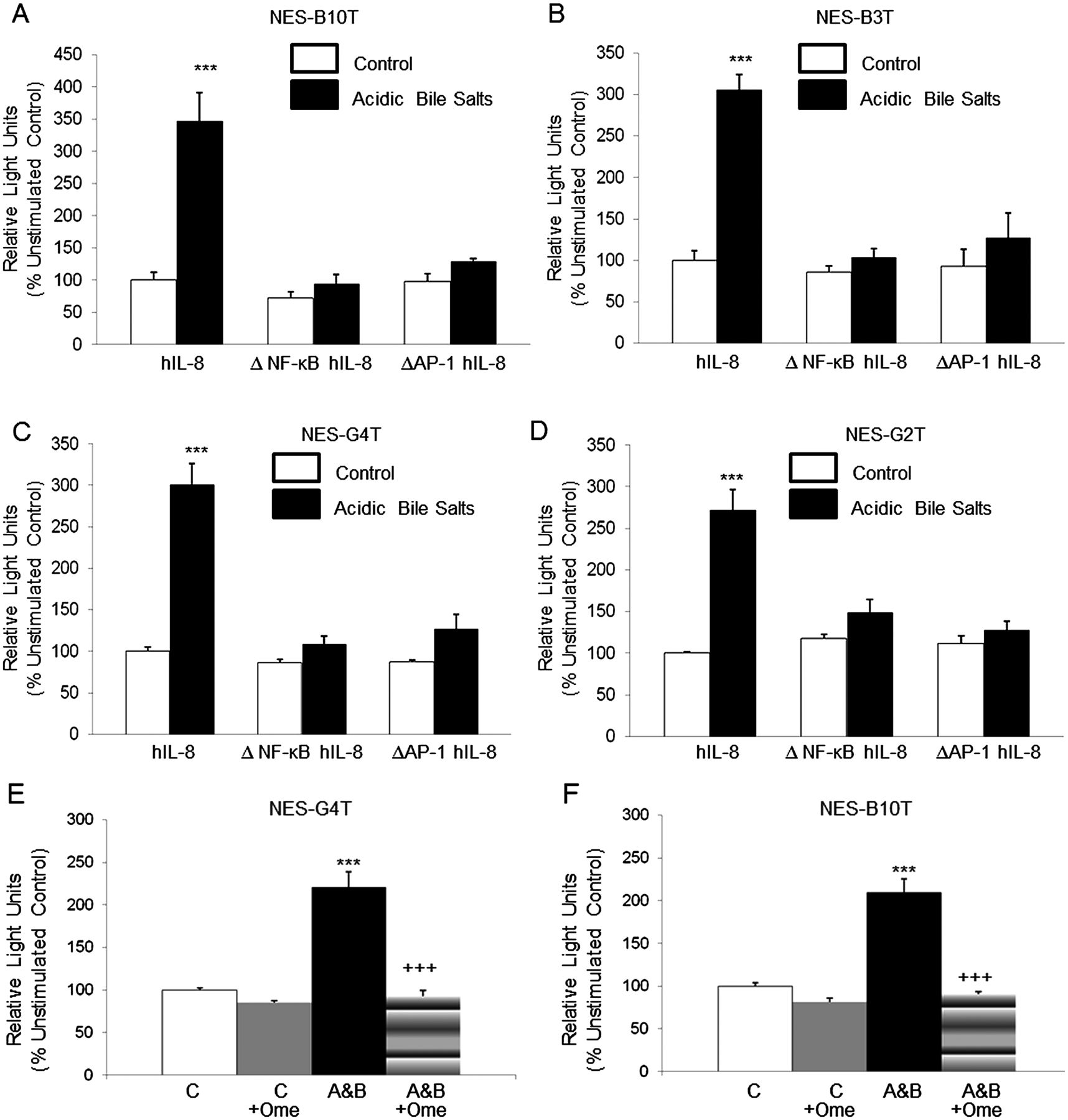
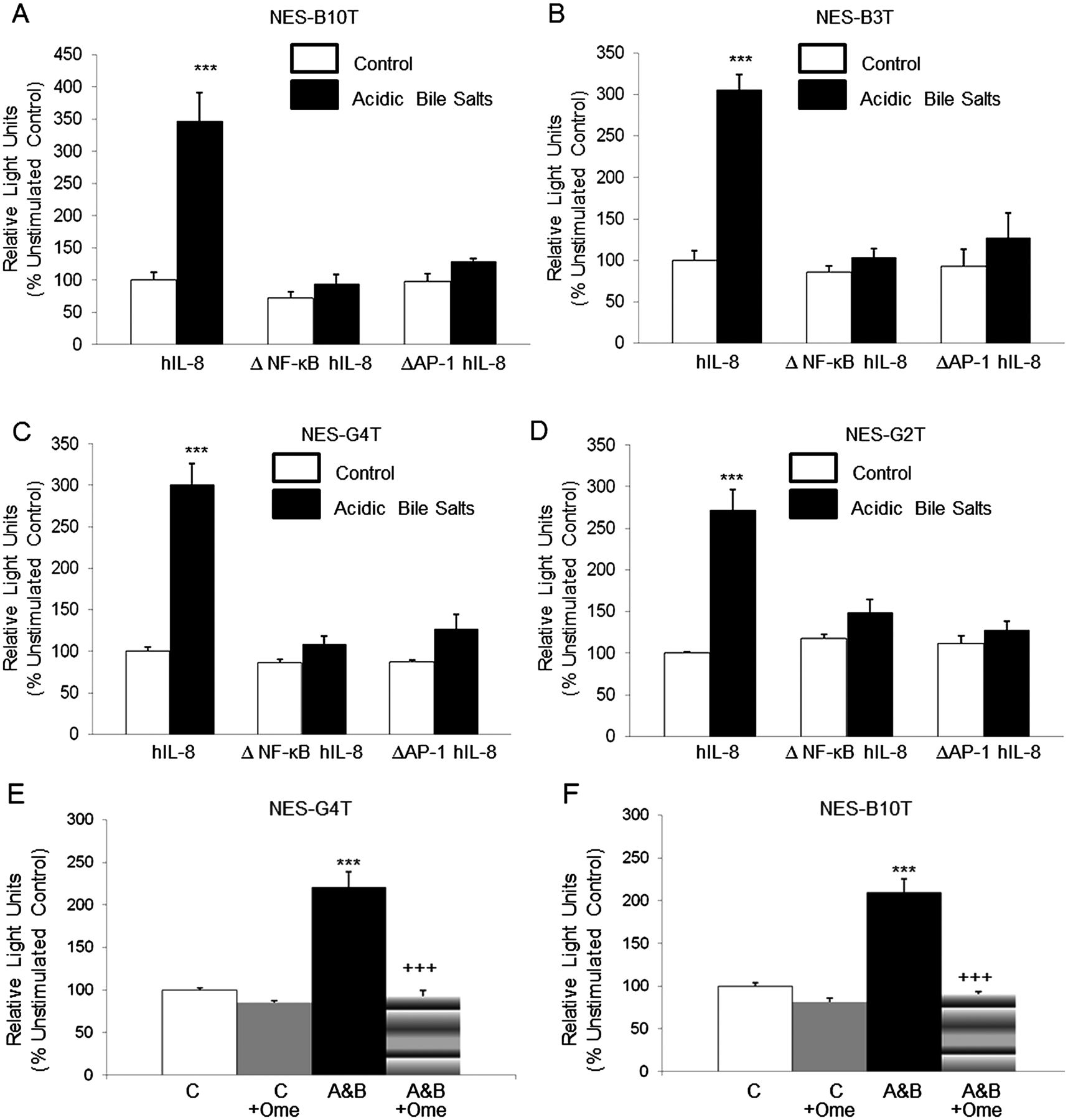
Acidic bile salt medium (A&B) increases IL-8 promoter activity by activating NF-κB and AP-1 binding sites in NES cell lines. (5A-D) Site-directed mutagenesis of the NF-κB binding site (ΔNF-κB hIL-8) or the AP-1 binding site (ΔAP-1 hIL-8) within the reporter construct pGL3-162/+44 hIL-8/Luc prevents activation of the IL-8 promoter in all four cell lines. (5E-F) Omeprazole (Ome) decreases activation of the IL-8 promoter by exposure to acidic bile salt media in (E) NES-G4T and (F) NES-B10T cells. The bar graphs depict the mean+SEM of at least two individual experiments. (***, p<0.001 compared with corresponding controls; +++, p<0.001 compared with corresponding acidic bile salt media-treated controls) RLU, relative light units.
The NF-κB and AP-1 DNA binding sites are necessary for acidic bile salt-stimulated IL-8 promoter activation in oesophageal squamous cell lines
A number of studies have demonstrated that the NF-κB DNA binding site in the IL-8 promoter is essential for transcriptional regulation of the gene (reviewed in ref. 20). The core IL-8 promoter also contains an activating protein (AP)-1 binding site that is not needed for transcriptional activation in some cells, but that contributes importantly to IL-8 promoter activation in others.20 We have previously shown that acidic bile salts induce signalling via the NF-κB and the mitogen-activated protein kinase (MAPK) pathways in oesophageal squamous cells.12 ,21 Therefore, we sought to determine the contribution of NF-κB and AP-1 transcription factors in the acidic bile salt-induced expression of IL-8. The −162/+44 fragment of the IL-8 promoter contains DNA binding sites for NF-κB and AP-1. To determine if the NF-κB and the AP-1 sites are necessary for IL-8 transcriptional activation in response to acidic bile salts, we transfected NES cell lines with ΔNF-κB 162/+44 hIL-8/Luc, which contains a point mutation in the NF-κB site, or with ΔAP-1 162/+44 hIL-8/Luc, which contains a point mutation in the AP-1 site. Point mutation of the NF-κB or AP-1 binding sites abolished IL-8 promoter activation in response to acidic bile salts (figure 6A–D). These findings suggest that NF-κB and AP-1 binding are essential for IL-8 promoter induction in response to acidic bile salts in oesophageal squamous cells.
Omeprazole decreases acidic bile salt-stimulated nuclear translocation of p65, and decreases the binding of p65, c-jun and c-fos to the IL-8 promoter in oesophageal squamous cell lines
Nuclear translocation and DNA binding are essential for transcription factors to activate the IL-8 promoter.20 Therefore, we explored whether omeprazole interfered with the ability of p65 (an NF-κB subunit), c-jun and c-fos (both AP-1 subunits) to undergo nuclear translocation and binding to the IL-8 promoter in acidic bile salt-stimulated NES cells. In untreated control cells, phospho-p65 and total p65 is found predominantly in the cytoplasm, with lesser amounts in the nucleus (figure 7A). ChIP assay demonstrates that the p65 protein binds to the IL-8 promoter (figure 7B). Omeprazole alone had minimal effects on baseline cytoplasmic and nuclear levels of phospho-p65 and total p65 (figure 7A). Stimulation with acidic bile salts decreased cytoplasmic levels of phospho-p65 and total p65, and markedly increased nuclear levels and binding of p65 to the IL-8 promoter; these effects were decreased markedly by treatment with omeprazole (figure 7A,B). In untreated control cells, c-jun and c-fos are located in the cytoplasm and nucleus, with baseline binding to the IL-8 promoter demonstrated by ChIP assay (figure 7A,B). Omeprazole alone had minimal effects on baseline cytoplasmic and nuclear expression of c-jun and c-fos (figure 7A). Stimulation with acidic bile salts decreased cytoplasmic levels of c-jun and c-fos, and increased the nuclear levels and binding of both these AP-1 subunits to the IL-8 promoter (figure 7A,B). Western blot revealed no differences in acidic bile salt-induced cytoplasmic and nuclear expression of c-jun and c-fos between omeprazole-treated and untreated NES cells (figure 7A). By contrast, omeprazole reduced binding of acidic bile salt-stimulated c-jun and c-fos to the IL-8 promoter to control levels (figure 7B). These findings suggest that omeprazole inhibits nuclear translocation of p65, but not c-jun or c-fos, and prevents binding of p65, c-jun and c-fos to the IL-8 promoter in oesophageal squamous cells.

Omeprazole (Ome) decreases acidic bile salt-induced (A&B) nuclear translocation of p65, but not of c-jun or c-fos in NES cells. Representative experiments of western blotting for (A) phospho-p65 and total p65 and c-jun and c-fos in cytoplasmic lysates and in nuclear lysates in NES-G4T and NES-B10T cells. Tubulin and TF-IID served as controls for the cytoplasmic and nuclear lysates, respectively. (B) Omeprazole (Ome) decreases acidic bile salt-induced p65, c-jun, and c-fos binding to the endogenous IL-8 promoter in NES-G4T and NES-B10T cells. Panels show representative experiments of chromatin immunoprecipitation (ChIP) assays for p65, c-jun and c-fos. Isotype-matched IgG served as a negative control. Numbers represent normalised densitometric values. C, untreated control cells.
Omeprazole treatment of oesophageal squamous cell lines exposed to acidic bile salts blocks the ability of the cells’ conditioned medium to induce immune cell migration
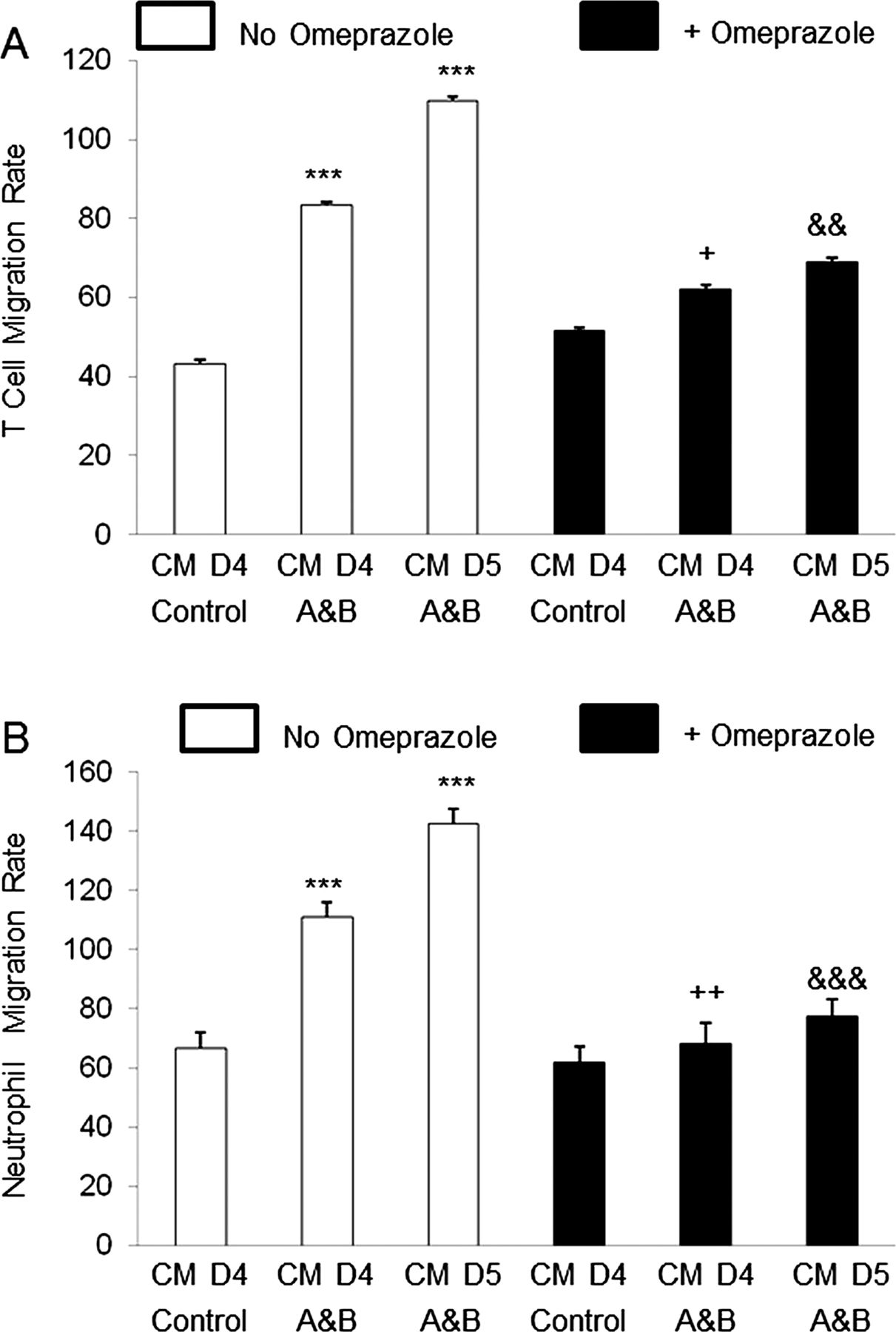
In earlier studies, we demonstrated that conditioned medium of oesophageal squamous cells treated with acidic bile salts induced migration of neutrophils and T cells, and that IL-8 in the conditioned medium contributed to that migration.11 In the present study, we found that omeprazole significantly decreased IL-8 secretion into conditioned medium of oesophageal squamous cells exposed to acidic bile salts. We also determined whether omeprazole treatment of NES cells exposed to acidic bile salts affected the ability of conditioned medium to induce immune cell migration. As in our earlier studies, we found that conditioned medium from NES-B10T cells exposed to acidic bile salts for 4 and 5 days significantly increased migration rates of T cells and neutrophils (figure 8A,B). Treatment with omeprazole significantly decreased the acidic bile salt-induced rise in immune cell migration rates (figure 8A,B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Migration rate of (A) T cells and (B) Neutrophils after exposure to conditioned medium (CM) collected from NES-B10T cells on day 4 in controls and on days 4 and 5 after exposure to acidic bile salt medium (A&B) in the presence or absence of omeprazole. The bar graphs depict the mean+SEM of at least 2 individual experiments. (***, p<0.001 compared with corresponding control; +, p<0.05 compared with corresponding CM day 4 A&B without omeprazole; ++, p<0.01 compared with corresponding CM day 4 A&B without omeprazole; &&, p<0.01 compared with corresponding CM day 5 A&B without omeprazole; &&&, p<0.001 compared with corresponding CM day 5 A&B without omeprazole).
Discussion
We have shown that exposure to acidic bile salt medium causes primary oesophageal squamous cells and telomerase-immortalised oesophageal squamous cell lines to secrete IL-8, a chemokine that is a major mediator of inflammation. We have found that omeprazole blocks this IL-8 secretion by suppressing activation of the IL-8 promoter, which decreases IL-8 mRNA expression stimulated by acidic bile salts. We have demonstrated that acidic bile salts activate the IL-8 promoter through NF-κB and AP-1 DNA binding sites, and that omeprazole blocks nuclear translocation of p65 (an NF-κB subunit) and decreases binding of p65, c-jun and c-fos (AP-1 subunits) to the IL-8 promoter. Finally, we have shown that omeprazole treatment of oesophageal squamous cells exposed to acidic bile salts blocks the ability of the cells’ conditioned medium to induce immune cell migration. These previously unknown PPI actions, which are entirely independent of effects on gastric acid secretion, might contribute to the beneficial effects of PPIs in the treatment of GORD. Further studies will be needed to establish that this acid-independent, anti-inflammatory effect can be achieved with the lower concentrations of omeprazole typically found in the blood of patients who take the drug.
Oesophageal mucosal biopsies from patients with reflux oesophagitis exhibit elevated levels of a number of proinflammatory cytokines in addition to IL-8 (eg, IL-1β, interferon-γ, IL-6, monocyte chemoattractant protein-1, RANTES),22–24 and it is possible that PPIs might interfere with the oesophageal expression of these other cytokines. We chose to focus on IL-8 (also called CXCL8)25 because it is known to be an important mediator of acute and chronic inflammation in a number of organs,26–28 and because IL-8 appears to play a key role in mediating reflux oesophagitis.11 ,29 Studies have documented IL-8 production by oesophageal epithelial cells in animal models of reflux oesophagitis,11 ,30 in human oesophageal cell lines,3 ,11 and in oesophageal mucosal biopsy specimens from patients with erosive and non-erosive reflux disease.23 ,31 ,32
IL-8 functions to attract and activate polymorphonuclear cells and lymphocytes, and oesophageal levels of IL-8 correlate with the severity of reflux oesophagitis.23 ,31 ,32 GORD treatment with PPIs causes a decrease in oesophageal mucosal IL-8 expression levels23 ,31 but it is not clear whether those levels decline merely because PPIs cause the healing of oesophagitis through their well-known antisecretory effects, or because PPIs exert direct suppressive effects on oesophageal IL-8 production. The observation that oesophageal mucosal IL-8 levels fall after successful antireflux surgery (without PPIs) suggests that the healing of oesophagitis alone can cause IL-8 levels to decline.24 Our study showing that PPIs can block reflux-induced activation of the IL-8 promoter in oesophageal epithelial cells suggests that this acid-independent effect of PPIs, along with their antisecretory effects, might contribute to the healing of reflux oesophagitis.
Proinflammatory stimuli known to increase IL-8 production also are known to activate protein kinases that modulate the activities of nuclear factor (NF)-κB and activator protein (AP)-1.20 These transcription factors regulate a number of key biological processes and, interestingly, NF-κB and AP-1 also appear to modulate each other's expression.33 Yoshida et al34 reported that human oesophageal epithelial cells transfected with luciferase expression vectors bearing mutations at the binding sites for either NF-κB or AP-1 exhibited suppression of bile salt-stimulated IL-8 gene transcription by 100% and 70%, respectively. Handa et al3 found that omeprazole and lansoprazole blocked the stimulated secretion of IL-8 by human gastric cancer and HUVEC cells, in part by preventing NF-κB activation.
To explore effects of acidic bile salts on IL-8 promoter activity, we transfected oesophageal cell lines using IL-8 promoters with or without mutations at the NF-κB or AP-1 binding sites. We confirmed that the acidic bile salt-induced increase in IL-8 promoter activity is dependent on NF-κB and AP-1 activation. We found that omeprazole blocked nuclear translocation of the p65 subunit of NF-κB, and reduced its binding to the IL-8 promoter. Omeprazole also interfered with binding of AP-1 subunits (c-jun and c-fos) to the IL-8 promoter. Taken together, these findings suggest that PPIs prevent acidic bile salt-induced, proinflammatory stimuli from increasing IL-8 secretion in oesophageal epithelial cells by blocking nuclear translocation of p65, and by preventing binding of p65, c-jun and c-fos to the IL-8 promoter. Gene promoter blocking effects of omeprazole are not unprecedented. In our recent studies using oesophageal epithelial cells from patients with eosinophilic oesophagitis, we found that omeprazole inhibited Th2 cytokine-stimulated secretion of eotaxin-3 by blocking STAT6 binding to the eotaxin-3 promoter through effects on chromatin remodelling.9
To explore possible functional effects of omeprazole-induced inhibition of chemokine production on the inflammatory response to gastro-oesophageal reflux, we used a double-chamber migration assay system to determine the ability of conditioned medium from oesophageal cells treated with acidic bile salts (with and without omeprazole pretreatment) to induce immune cell migration. We found that conditioned medium from oesophageal squamous cells exposed to acidic bile salts caused a significant increase in migration of neutrophils and T cells, and that this increased immune cell migration was blocked by treating the oesophageal squamous cells with omeprazole. It is not clear whether this resulted solely from the omeprazole-induced decrease in secretion of IL-8 by the oesophageal squamous cells, or from effects of omeprazole on cell secretion of other chemokines.
In conclusion, we have shown that omeprazole inhibits IL-8 secretion stimulated by exposure to acid and bile salts in oesophageal epithelial cells. We have demonstrated that acidic bile salts activate the IL-8 promoter through NF-κB and AP-1 DNA binding sites, and that omeprazole inhibits IL-8 production by blocking nuclear translocation of p65 and by decreasing binding of p65, c-jun and c-fos to the IL-8 promoter. All these omeprazole effects are entirely independent of effects on gastric acid secretion. Thus, we have demonstrated a novel mechanism that might contribute to the beneficial effects of PPIs in the healing of GORD.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
Contributors All authors included in this paper fulfil the criteria of authorship and have approved the submission of this manuscript. Specific contributions are as follows. XH: study design; technical and material support; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript. XZ: study design; technical and material support; analysis and interpretation of data; important intellectual content. CY: technical and material support; important intellectual content. QZ: technical and material support; important intellectual content. EC: technical and material support; important intellectual content. DHW: technical and material support; important intellectual content. THP: technical and material support; important intellectual content. SJS: study concept; analysis and interpretation of data; critical revision of manuscript; important intellectual content. RFS: study concept/design; analysis and interpretation of data; critical revision of manuscript; important intellectual content; drafting of manuscript.
-
Funding This work was supported by the Office of Medical Research, Departments of Veterans Affairs (R.F.S., S.J.S., D.H.W.), VISN 17 Start-Up Award (Q.Z.), the National Institutes of Health (R01-DK63621 to R.F.S., R01-CA134571 to R.F.S. and S.J.S., K12 HD-068369-01 to E.C.), the American Gastroenterological Association Institute (Fellow to Faculty Transition Award to E.C.) and NASPGHAN Foundation/AstraZeneca Award (E.C.).
-
Competing interests None.
-
Ethics approval Institutional Review Board of the Dallas VA Medical Center.
-
Provenance and peer review Not commissioned; externally peer reviewed.