Article Text

Abstract
Objective The complex genetic aetiology underlying irritable bowel syndrome (IBS) needs to be assessed in large-scale genetic studies. Two independent IBS cohorts were genotyped to assess whether genetic variability in immune, neuronal and barrier integrity genes is associated with IBS.
Design 384 single nucleotide polymorphisms (SNPs) covering 270 genes were genotyped in an exploratory cohort (935 IBS patients, 639 controls). 33 SNPs with Puncorrected<0.05 were validated in an independent set of 497 patients and 887 controls. Genotype distributions of single SNPs were assessed using an additive genetic model in IBS and clinical subtypes, IBS-C and IBS-D, both in individual and combined cohorts. Trait anxiety (N=614 patients, 533 controls), lifetime depression (N=654 patients, 533 controls) and mRNA expression in rectal biopsies (N=22 patients, 29 controls) were correlated with SNP genotypes.
Results Two SNPs associated independently in the exploratory and validation cohort: rs17837965-CDC42 with IBS-C (ORexploratory=1.59 (1.05 to 1.76); ORvalidation=1.76 (1.03 to 3.01)) and rs2349775-NXPH1 with IBS-D (ORexploratory=1.28 (1.06 to 1.56); ORvalidation=1.42 (1.08 to 1.88)). When combining both cohorts, the association of rs2349775 withstood post hoc correction for multiple testing in the IBS-D subgroup. Additionally, three SNPs in immune-related genes (rs1464510-LPP, rs1881457-IL13, rs2104286-IL2RA), one SNP in a neuronal gene (rs2349775-NXPH1) and two SNPs in epithelial genes (rs245051-SLC26A2, rs17837965-CDC42) were weakly associated with total-IBS (Puncorrected<0.05). At the functional level, rs1881457 increased IL13 mRNA levels, whereas anxiety and depression scores did not correlate with rs2349775-NXPH1.
Conclusions Rs2349775 (NXPH1) and rs17837965 (CDC42) were associated with IBS-D and IBS-C, respectively, in two independent cohorts. Further studies are warranted to validate our findings and to determine the mechanisms underlying IBS pathophysiology.
- GENETIC POLYMORPHISMS
- IMMUNE RESPONSE
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
Previous studies showed that irritable bowel syndrome (IBS) exhibits the typical features of a complex multifactorial disorder influenced by both environmental and genetic factors.
-
Defects in mucosal barrier integrity, inflammation and immune interactions with bacteria in the gut, neuronal sensitisation (altered nociceptor expression, increased anxiety and depression) and altered motor function are emerging as candidate genes contributing to the pathogenesis of IBS.
-
Although TNFSF15 variants have repeatedly been associated with IBS, genetic studies in IBS have been largely inconclusive due to small sample sizes.
What are the new findings?
-
Two single nucleotide polymorphisms, rs17837965 in CDC42 and rs2349775 in NXPH1, were associated with IBS-C and IBS-D, respectively, in two independent cohorts of 1432 IBS patients and 1526 controls.
-
The rs2349775 variant in NXPH1 withstood post hoc correction for multiple testing in the combined IBS-D subgroup suggesting that it represents a true susceptibility factor for IBS.
How might it impact on clinical practice in the foreseeable future?
-
Our data may contribute to the identification of important targets in IBS which may ultimately result in improved clinical management.
Background
Irritable bowel syndrome (IBS) is a multifactorial disorder, which is more common in women and presents with a variety of symptoms, including altered bowel habits, bloating and increased abdominal pain sensation.1 As IBS diagnosis is based on symptoms, it consists of several clinical subtypes including constipation predominant IBS (IBS-C), diarrhoea predominant IBS (IBS-D) or alternating constipation and diarrhoea IBS (IBS-A) with potentially different underlying pathophysiological pathways. Diet, psychological distress, infections, low-grade inflammation and immunological alterations, dysregulated interactions with potentially altered microbiota, sensitisation/upregulation of nociceptors and changes in intestinal motility and altered gut–brain communication have all been proposed as contributing factors.2–4
Epidemiological studies of familial aggregation5 ,6 and twins7–11 suggest that genetic predisposition may explain up to 22%–48% of the incidence of IBS. However, these data are conflicting as Mohammed et al demonstrated that social learning has an equal or greater influence.12 Moreover, environmental risk factors such as psychological stress, smoking and the use of antibiotics8 significantly contribute to IBS susceptibility as well. Overall, IBS exhibits typical features of a complex genetic disorder influenced by both environmental and genetic factors. Although our current knowledge on genetic predisposition to IBS is very limited, a number of studies already assessed the risk effects of single nucleotide polymorphisms (SNPs) in candidate genes. One study reported that SNPs in TLR9, IL6 and CDH1, which are involved in immune activation and epithelial barrier integrity, were nominally associated with postinfectious IBS (PI-IBS).13 Another study focused on functional gastrointestinal disorders and correlated colonic transit rate and pain sensation with polymorphisms in the neuropeptide S receptor gene (NPSR1),14 a gene involved in inflammation, anxiety and nociception. In the largest genetic study of IBS so far, a strong association was reported between rs4263839 in TNFSF15,15 which is involved in Th17 immune response, and IBS (particularly IBS-C) in two independent cohorts from Sweden and USA. Although with a different subtype (IBS-D) and with another SNP, this association was recently replicated in UK individuals.16 Additional smaller studies identified SNPs in serotonergic genes17–21 and inflammatory genes22–24 as susceptibility SNPs for IBS. All genetic variants previously associated with IBS have been reviewed elsewhere.25–27
In view of the diversity of IBS aetiologies and phenotypic subtypes (IBS-A, IBS-C, IBS-D, PI-IBS), there is a great need for much larger studies systematically evaluating genetic variants in IBS. We therefore evaluated genetic predisposition to IBS, using a hypothesis-driven genetic association approach with validation in an independent cohort (except for the genes that were already reported in this validation cohort). In particular, 384 variants located in 270 genes involved in immune modulation, inflammation, motor function, pain processing, nociceptor sensitisation, altered neuronal function (centrally and peripherally) and epithelial barrier integrity were genotyped in a cohort of 1432 IBS patients.
Subjects and methods
Study populations
Two independent cohorts of IBS patients and controls from UK/US/Canada and Sweden were included in this study. Informed consent was obtained from all participants and local ethics committees approved the study protocol. The demographics and clinical characteristics of both cohorts are reported in table 1. The IBS patients were divided into subgroups based on the predominant bowel habit according to the Rome II criteria.1
Demographic and clinical characteristics of IBS patients and controls
Exploratory cohort: This cohort consisted of 935 IBS patients and 639 controls from the UK, USA or Canada. All patients confirmed Caucasian ethnicity. IBS patients were included if they had a history of IBS for at least 6 months according to Rome II criteria. IBS patients had a colonoscopy/barium enema during the last 12 months with normal results, supporting an IBS diagnosis. Controls were included if they had no previous IBS diagnosis (based on physician assessment and negative Rome II criteria) and no former history of chronic gastrointestinal diseases. Controls were matched to IBS patients according to age (±5 years), gender, ethnicity and referring physician, where feasible. All samples from the exploratory cohort were kindly provided by GlaxoSmithKline (UK).
Validation cohort: A total of 497 IBS patients were recruited from primary, secondary and tertiary care centres and clinics in Stockholm, Gothenburg, Malmoe and Kalix-Haparanda in Sweden. The diagnosis of IBS was based on Rome II criteria and symptom phenotypes, consensus criteria and validated questionnaires of gastrointestinal and somatic symptoms. In all, 887 controls were healthy blood donors free of gastrointestinal presentations and inflammatory disease.
Genotyping
To select genetic variants in genes involved in immune modulation, inflammation, motor function, pain processing, nociceptor sensitisation and altered neuronal function (centrally and peripherally), we performed a systematic review using the public database PubMed: with highest priority, we included SNPs previously characterised and potentially associated with IBS in serotonergic pathways, immune responses, and intestinal epithelial barrier and motor function13–15 17–22 28–30 (keywords ‘Irritable bowel syndrome’ and ‘SNP’ or ‘genetic variant’). Next, using the same overall approach, we selected SNPs that were previously identified as susceptibility variants in independent association studies of diseases with potentially comparable underlying pathophysiological mechanisms (keywords were ‘SNP’ or ‘genetic variant’ and the following diseases: diseases mediated by aberrant immune activation and/or inflammation: Crohn's disease, ulcerative colitis, type 1 diabetes, rheumatoid arthritis, multiple sclerosis, chronic obstructive pulmonary disease, ankylosing spondylitis, hyper IgE syndrome, barrier dysfunction related diseases: coeliac disease and diseases due to altered neuronal function: anxiety, major depression, somatic pain sensation). Finally, we also selected SNPs based on a HAPMAP tagging approach (http://www.hapmap.org) in tight-junction genes, nociceptors such as TRP genes, Toll-like receptors, NOD-like receptors and cytokines (see list of selected SNPs in online supplementary table S1). In total, 443 SNPs were selected, evaluated and scored by the Illumina Technical Support Service for potential genotyping using Illumina Golden Gate. SNPs with a GGGT score lower than 0.6 were excluded. Likewise, we excluded tagging SNPs if more than four tagging SNPs per gene were selected. Of the remaining 384 SNPs, 318 SNPs had a GGGT score above 0.8 and 66 SNPs had a GGGT score between 0.6 and 0.8.
The final 384 SNPs included 25 SNPs that were previously associated with IBS,13–15 17–22 28–31 265 SNPs that were previously identified as susceptibility variants for immune mediated diseases, genes involved in barrier dysfunction related diseases and neuropathologies and 94 SNPs were selected based on the SNP tagging approach. Out of the 384 SNPs, 19 SNPs had a minor allele frequency (MAF) <0.05 in the Utah residents with ancestry from northern and western Europe (CEPH) population (Utah residents with Northern and Western European Ancestry, CEU).
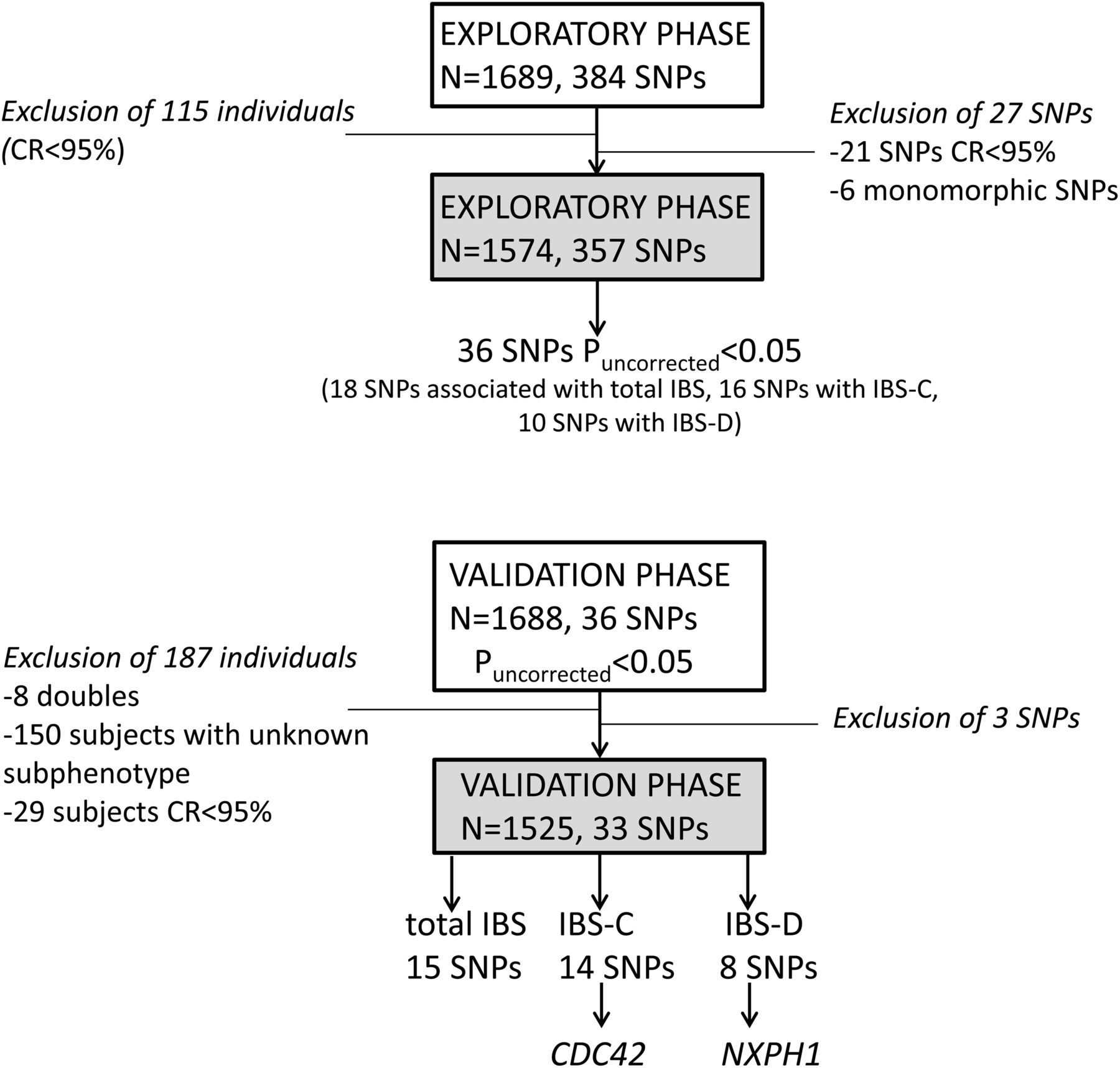
Samples from the exploratory cohort were genotyped with custom-designed oligo chips using Illumina Golden Gate (Vesalius Research Centre, KULeuven). Based on the results of this exploratory study, SNPs nominally associated with total IBS, IBS-C or IBS-D with Puncorrected<0.05 were selected for validation in the Swedish cohort by Sequenom MassARRAY (figure 1). Quality control criteria for Golden Gate and Sequenom MassARRAY genotyping included: a SNP call rate of >0.95 in patients and controls, a sample call rate >0.95, an MAF of >0.01 and a Hardy–Weinberg equilibrium p value of >0.0001 in controls and >0.00001 in cases.

Study design.
The State-Trait Anxiety Inventory anxiety and depression questionnaires and the mRNA expression analysis in colonic biopsies of the exploratory cohort are described in online supplementary material and methods.
Statistical analyses
Fisher’s exact tests were used to test for Hardy–Weinberg equilibrium. For the analysis of individual SNPs, we used the Cochran–Armitage trend test under the assumption of an additive genetic model and Plink V.1.07 software (http://pngu.mgh.harvard.edu/~purcell/plink/). Furthermore, we performed a Cochran–Mantel–Haenszel meta-analysis in the combined cohort using the Statistical Analysis Software (http://www.sas.com). Analyses were performed for all IBS patients and two IBS subgroups (IBS-C and IBS-D). The 140 IBS-A samples in our exploratory cohort did not provide enough power to detect statistical significance. SNPs with p<0.05 in the IBS-A subgroup were therefore not further validated. SNPs with a Puncorrected<0.05 in the exploratory cohort were selected for follow-up analysis in the validation cohort. To correct for multiple testing (33 SNPs in three IBS groups, ie, total IBS, IBS-C and IBS-D), a p value <5E-04 was considered significant. In addition, False Discovery Rate correction was applied for the number of SNPs per IBS subgroup (figures 2⇓–4, see online supplementary tables S1–S3) (R project software V.2.15; http://www.r-project.org/). Linkage disequilibrium between markers in the two studied populations was based on HapMap data using Haploview V.4.2 software (http://www.broadinstitute.org/haploview). The CaTS power calculator for two-stage association studies was used for power calculations. RT-qPCR results, trait anxiety and depression scores were analysed with Graphpad Prism software (GraphPad Software, Inc., La Jolla, California, USA) using the Mann–Whitney U test for unpaired samples.

Association of genetic variants with total irritable bowel syndrome (IBS). Shown from left to right are: Nearest gene with National Center for Biotechnology Information (NCBI) dbSNP database accession numbers (rs number) in the respective cohort (exploratory cohort (UK), validation cohort (SWE), and combined cohort (COMB), Armitage's Trend test (P_Trend), Cochran–Mantel–Haenszel meta-analysis (P_CMH), false positive report P_CMH (FDR P_CMH), allelic OR and lower and upper bound of the corresponding 95% CI). *SNP was not associated with total IBS but with C-IBS in the exploratory cohort. FDR, False Discovery Rate; SNP, single nucleotide polymorphism.

Association results of genetic variants with constipation predominant irritable bowel syndrome. Shown from left to right are: Nearest gene with National Center for Biotechnology Information (NCBI) dbSNP database accession numbers (rs number) in the respective cohort (exploratory cohort (UK), validation cohort (SWE), and combined cohort (COMB), Armitage's Trend test (P_Trend), Cochran–Mantel–Haenszel meta-analysis (P_CMH), false positive report P_CMH (FDR P_CMH), allelic OR and lower and upper bound of the corresponding 95% CI). FDR, False Discovery Rate; SNP, single nucleotide polymorphism.
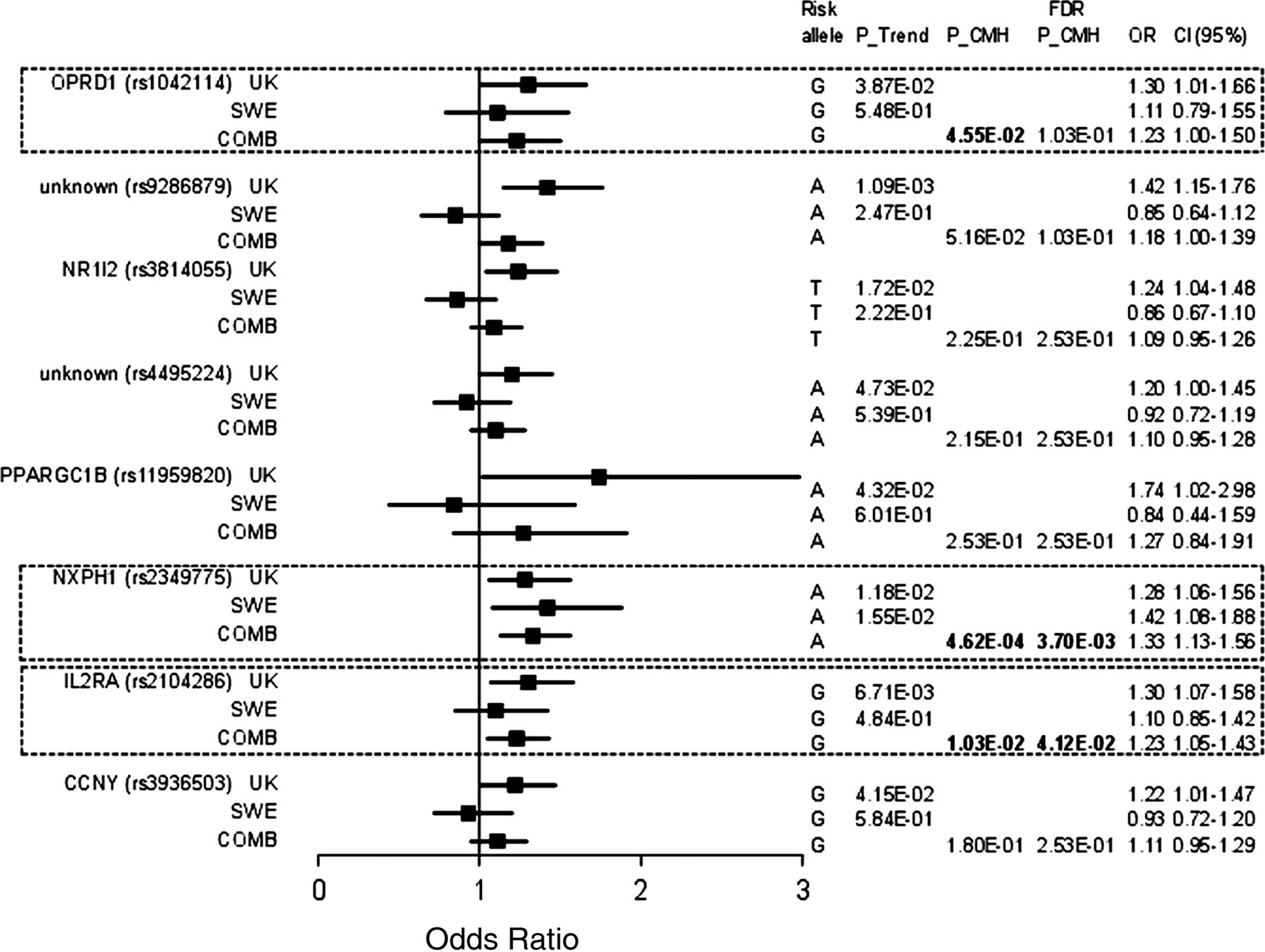
Association results of genetic variants with diarrhoea predominant irritable bowel syndrome. Shown from left to right are: Nearest gene with National Center for Biotechnology Information (NCBI) dbSNP database accession numbers (rs number) in the respective cohort (exploratory cohort (UK), validation cohort (SWE), and combined cohort (COMB), Armitage's Trend test (P_Trend), Cochran–Mantel–Haenszel meta-analysis (P_CMH), false positive report P_CMH (FDR P_CMH), allelic OR and lower and upper bound of the corresponding 95% CI). FDR, False Discovery Rate; SNP, single nucleotide polymorphism.
Results
Genetic variants associated with IBS
Out of 384 SNPs, 21 SNPs failed our quality control due to low call rates, while another six SNPs were excluded because they were monomorphic. All remaining 357 SNPs were subsequently assessed for association with IBS and its different clinical subtypes (935 IBS patients, of whom 430 had IBS-D and 325 had IBS-C, and 639 controls) in our exploratory cohort. Genotype distributions and association results of all SNPs are reported in online supplementary tables S4–S7.
Total IBS
In total, 18 SNPs were weakly associated (Puncorrected<0.05) with total IBS in the discovery cohort (see online supplementary table S5). The rs744166 variant in STAT3 was previously reported not to be associated with IBS in the validation cohort15 ,28 and was therefore not replicated. Overall, three SNPs failed genotyping in the validation cohort: rs1758241 (unknown gene) was not further studied, rs3024505 (IL10) and rs1800925 (IL13) failed but nearby located SNPs rs1800896 (R2=0.253) and rs1881457 (R2=0.747) respectively—also identified in the exploratory cohort—were successfully genotyped. Only one SNP, rs4986791 near TLR4, was weakly associated (Puncorrected<0.05) with IBS in the exploratory and validation cohorts, but this SNP had opposite allelic effects in the two cohorts (ORExploratory=3.38 (1.53 to 7.44) and ORvalidation=0.47 (0.31 to 0.7)) (figure 2, see online supplementary table S1). Next, we combined the exploratory and validation cohorts and we assessed using a Cochran–Mantel–Haenszel meta-analysis whether SNPs associated significantly with total IBS (figure 2, see online supplementary table S1). Although none of the validated SNPs was resistant for multiple testing (p<5E-04), six SNPs were weakly associated at the uncorrected threshold (Puncorrected<0.05), namely, rs1464510 in Lipoma-preferred partner (LPP), rs1881457 in IL13, rs245051 in SLC26A2, rs2349775 in NXPH1, rs2104286 in IL2RA and rs17837965 near CDC42 (figure 2, see online supplementary table S1 and online supplementary figure S1).
IBS subphenotypes
All SNPs with a Puncorrected<0.05 in the IBS-C or IBS-D subgroup of the exploratory cohort were selected for validation. One SNP, namely rs17837965 in CDC42, was associated with IBS-C in both independent cohorts (PCMH IBS-C=2.88E-03; ORexpl=1.59 (1.05 to 1.76) and ORvalidation=1.76 (1.03 to 3.01)) (figure 3, see online supplementary table S2) and may represent a true association signal. Also for IBS-D, only one SNP, namely SNP rs2349775 (NXPH1), was associated with IBS-D in both cohorts. Moreover, rs2349775 was the only SNP that withstood post hoc correction for multiple testing (PCMH IBS-D=4.62E-04; ORexpl=1.28 (1.06 to 1.56) and ORvalidation=1.42 (1.08 to 1.88)) (figure 4, see online supplementary table S3) and may therefore represent a true association signal.
Functional analysis of the newly identified candidate SNPs
Next, in an attempt to provide functional evidence that the six SNPs weakly associated with total IBS (Puncorrected<0.05) were functionally implicated in IBS pathophysiology, we correlated SNP genotypes with expression of their target genes. Because the function of SNP rs2104286 in IL2RA has already been reported in literature (ie, rs2104286 modulates soluble IL2RA serum levels in healthy controls, multiple sclerosis and type 1 diabetes patients32), these SNPs was not studied at the functional level. The neuronal gene NXPH1 was not detected by qPCR in rectal biopsies or PBMCs of neither IBS nor controls (n=3, data not shown). This SNP was therefore correlated with IBS comorbidity symptoms, that is, anxiety and depression.
Association of NXPH1 with anxiety and depression
NXPH1 (rs2349775) was previously associated with neuroticism33 and was preferentially associated with IBS-D in our study. Trait anxiety was slightly higher in IBS compared with controls (IBS (n=614): 45 (IQR 42–48) vs controls (n=533): 44 (IQR 42–47), p=0.0002). There was no correlation between the rs2349775 (NXPH1) genotype and trait anxiety (data not shown). Besides anxiety, IBS patients had higher life time depression scores compared with controls (IBS (n=654): 7, IQR 0–9 vs controls (n=533): 3 (IQR 0–8), p<0.0001). However, in IBS and controls, depression scores were not affected by the genotype of rs2349775 (NXPH1) (GG (n=79): 4 (IQR 0–9); AG (n=448): 6 (IQR 0–9); AA (n=586): 6 (IQR 0–9)).
Functional effect of rs1881457 on IL13 mRNA expression in rectal biopsies
The rs1881457 SNP in IL13 is located on chromosome 5q31 in a genomic region containing a cluster of Th2 cytokine genes (IL13, IL4 and IL5). IL13 mRNA expression was quantified in rectal biopsies of 22 IBS patients and 29 controls where we compared the extreme phenotypes, wild type versus homozygous risk allele carriers. IL13 mRNA expression was significantly higher in the rs1881457 AA than in the CC genotype carriers (PMann Whitney U<0.0001; figure 5A). This effect was seen in IBS patients (PMann Whitney U=0.048; figure 5B) and controls (PMann Whitney U=0.0004; figure. 5B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
IL13 mRNA expression is dependent on the rs1881457 genotype. IL13 mRNA expression was decreased in the rs1881457 CC genotype compared with the AA genotype (A). In both irritable bowel syndrome (IBS) and controls, the protective C alleles resulted in decreased IL13 mRNA expression compared with the A risk alleles (B). There was no difference in IL13 mRNA expression in controls (ctrls) compared with IBS (B). Each dot represents an individual; the line represents the median.
Expression profiles in rectal biopsies of nominally significant SNPs in epithelial genes
Although rs245051 in SLC26A2 is also located on chromosome 5q31, this SNP occurs independently of rs1881457 (IL13) (r2=0). There was no difference in SLC26A2 mRNA expression between IBS patients and controls (data not shown). Moreover, rs245051 did not affect SLC26A2 mRNA levels in IBS (data not shown). Finally, we were not able to assess the effect of rs17837965 on CDC42 mRNA expression due to a lack of biopsies exhibiting the less frequent GG genotype.
Discussion
We demonstrated that genetic variants in CDC42 and NXPH1 were associated with IBS-C and IBS-D, respectively, in two independent cohorts. Only one SNP, namely rs2349775 (NXPH1), withstood post hoc correction for multiple testing. Moreover, six novel potential candidate risk loci were weakly associated with IBS (Puncorrected<0.05) in the combined cohort, namely, three SNPs in immune related genes IL2RA, IL13 and LPP, one SNP in the neuronal expressed gene NXPH1, and two SNPs in epithelial expressed genes SLC26A2 and CDC42.
SNPs located in immune-related genes
Immune activation or dysregulation has been proposed as underlying mechanism of IBS, especially following an episode of infectious gastroenteritis. Besides increased numbers of T cells and enteroendocrine cells in PI-IBS rectal biopsies,34 much attention has been given to the role of mast cells, monocytes and T cells in other IBS subtypes.2 We potentially identified three novel SNPs associated with IBS in genes with an immune function, namely, rs2104286 in interleukin two receptor α (IL2RA), rs1464510 in LPP and rs1881457 in interleukin 13 (IL13).
Rs1881457 in IL13 is located at chromosome 5q31 in the promotor region of IL13, a cluster of genes involved in Th2 immune response. Kiesler et al reported that the rs1881457 C allele, which is associated with allergic asthma, results in enhanced IL13 expression in vitro.35 In contrast, we found that the A allele was weakly associated with IBS and rs1881457*AA biopsies expressed more IL13 mRNA compared with rs1881457*CC biopsies, an effect seen in both IBS patients and controls. The A risk allele was present in 82% and 80% of IBS patients compared with 79% and 77% of controls in the exploratory and validation cohort, respectively. Our cohort provided 62% power to detect a positive association (Puncorrected<0,05) for rs1881457 (supplementary table 9). Altogether, our results indicate that IBS patients carry more A alleles compared with healthy individuals and carriers of this allele express more IL13 mRNA compared with C allele carriers. Because rs1881457 is located in a dense, highly correlated region, we cannot deduct which SNP is causal or reflects other causal genetic variants. Nevertheless, we speculate that in a minority of IBS patients, genotypic variants in the Th2 cytokine cluster at 5q31 may contribute to IBS pathogenesis by increasing IL13 expression, possibly even more pronounced in response to pathogens or intraluminal antigens.
Of note, although, in yet another subset of patients (IBS-A) compared with the original observations (all IBS subtypes and IBS-C,15 or IBS-D16), we detected association with the TNFSF15 gene in the exploratory cohort (for SNP rs4263839: Ptrend=4.26E-02; SNP rs6478108: Ptrend=4.09E-02, online supplementary table S8). All IBS-A patients in the exploratory cohort derived from Canada or US sites and did not include UK samples;16 hence this is truly a repeat. Genetic variation at this locus has now been repeatedly associated with IBS and its subtypes in four different case-control cohorts, namely, from Sweden,15 USA,15 UK16 and now in a US/Canada cohort, which strongly suggests this may be in fact the first true causative locus identified in IBS.
SNPs in genes involved in neuronal functioning
IBS patients have more psychopathology, particularly anxiety disorders and depression, compared with the general population.36 Rs2349775 in NXPH1 was previously associated with neuroticism.33 In two independent cohorts, rs2349775 in NXPH1 was associated with IBS-D (75% of patients vs 70% of controls in exploratory cohort, 77% of patients vs 70% of controls in validation cohort). This is the only SNP that withstood correction for multiple testing. In the IBS-D cohort, we had 79% statistical power to detect a positive association for this SNP (Puncorrected<0.05). In line with the available literature, lifetime depression37 was increased in our exploratory IBS cohort compared with controls. However, rs2349775 in NXPH1 had no effect on depression or trait anxiety levels. Besides having central effects, secreted NXPH1 inhibits the proliferation of haematopoietic progenitor cells involved in the immune response.38 We speculate that genetic variants in NXPH1 may alter the immune response to a gastrointestinal insult, resulting in low grade inflammation and diarrhoea. However, the functional role of genetic variations in NXPH1 on IBS susceptibility and diarrhoea needs to be further explored.
SNPs in epithelial genes
The epithelial lining of the intestine is crucial in maintaining tolerance and controls antigen exposure to the mucosal immune system. One of the mechanisms proposed to contribute to the pathophysiology of IBS is impaired intestinal barrier function.39 In the present study, we found a consistent association between rs17837965 in cell division cycle 42 (CDC42) and IBS-C in two independent cohorts (G allele in 6.5% patients vs 4.2% controls in exploratory cohort, 7.2% of patients vs 4.2% controls in validation cohort). CDC42 is associated with schizophrenia,40 is crucial for normal dendritic spine development in the brain40 and is also involved in intestinal stem cell differentiation and proliferation.41 Intestinal stem cells lacking CDC42 undergo defective cell division, abnormal morphogenesis, elevated apoptosis and failed Paneth cell differentiation,41 ,42 indicating that genetic variants in CDC42 may alter epithelial barrier function. Moreover, the loss of absorptive microvilli from the surface of small intestinal enterocytes (Caco-2 cells) in response to Escherichia coli or its toxins was linked to CDC42 functionality.43 Additionally, CDC42 regulates epithelial membrane transport and secretion.41 ,44 Altogether, we speculate that genetic variants of CDC42 may alter secretion and/or barrier function, resulting in constipation. Rs17837965 GG carriers are rather rare (4%–7%) making it difficult to perform functional studies aiming to dissect the functional effect of genetic variants in CDC42.
Finally, we identified a weak association between SLC26A2 and IBS(-D). SLC26a2 is an ubiquitously expressed electroneutral SO(4)(2-) transporter with high expression levels in colonic epithelial lining.45 ,46 SLC26A2 is downregulated in Crohn's disease47 but there was no difference in SLC26A2 mRNA expression between IBS and controls. Again, little is known about SLC26A2 and how genetic variants of SLC26A2 contribute to IBS remains to be evaluated.
In summary, our data indicate that genetic variants in genes involved in neuronal processing (NXPH1), and epithelial barrier function (CDC42) might contribute to IBS pathogenesis. However, although our results are encouraging, it needs to be stressed that the SNPs identified in our study are weakly associated (Puncorrected<0.05) and despite biological plausibility may represent false positive associations. Hence, our findings are hypothesis-generating and still need to be confirmed in independent studies. We would like to emphasise, however, that they were identified in a much larger IBS population than normally is the case in the IBS field. We therefore believe that our studies may serve as a very valuable source to select and confirm candidate susceptibility SNPs or genes involved in IBS.
Nevertheless, our study is limited by the fact that SNPs were often selected based on genome wide association studies focusing on diseases with potentially comparable underlying pathophysiological mechanisms as IBS. Therefore, our selection contained several intronic SNPs as well as SNPs with an unknown function or SNPs with an (yet) unclear contribution to IBS pathogenesis. Our current sample size provides 79% power to detect associations with a p value <5E-4 and a risk effect size of 1.25 and MAF of 0.40 (see online supplementary table S10). To reach genome-wide significance (α=10E-7), a two-stage approach with 5000 IBS patients and 5000 healthy controls would, for instance, be required to have 95% power to detect associations with SNPs having an MAF of 0.20 and risk of 1.25. Such high number of patients was not available, and therefore an extensive hypothesis driven genetic association approach was used instead. As a result, none of the selected SNPs could be used as population-based markers to assess potential population stratification. Additionally, IBS diagnosis is very common (15%) in the general population; IBS risk variants may therefore also be present in a significant proportion of controls. However, since we used controls that are free of IBS diagnosis, our results could sustain this bias and may become even more significant when comparing IBS patients with an ideal set of controls, which would consist of healthy volunteers devoid of any of the IBS inclusion criteria. Finally, in the future, the discovery of IBS disease genes may be complemented by studies focusing on endophenotypes underlying IBS, such as colonic transit and visceral pain.
In conclusion, genetic variants in CDC42 and NXPH1 were associated with IBS-C and IBS-D respectively in two independent cohorts. Further studies are warranted to validate our findings and to determine the mechanisms by which these variants underlie IBS pathophysiology.
Acknowledgments
The authors would like to thank Dr Theadore Ptak, Renee Henry, Ellen Goldstein, Cindy Lee, Deborah Roach, Jacqueline Rabuzin and Elizabeth Crosland (Toronto Digestive Disease Associates (TDDA Inc.), Toronto, Ontario, Canada), Dr Mark Silverberg and Lori Baladjay (Mount Sinai Hospital, Toronto, Ontario, Canada), Dr Yehuda Ringel, Dr Robert Sandler, Alesia N Aileo, Sarah Causey and Sarah Yeskel (University of North Carolina at Chapel Hill, North Carolina, USA) for collecting samples and acquiring data and Dr Rachel Gibson (GlaxoSmithKline) for logistic support. We thank all of the patients and the supporting staff at each site.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure
- Data supplement 10 - Online table 8
- Data supplement 11 - Online table 9
- Data supplement 12 - Online table 10
- Data supplement 2 - Online methodology
- Data supplement 3 - Online table 1
- Data supplement 4 - Online table 2
- Data supplement 5 - Online table 3
- Data supplement 6 - Online table 4
- Data supplement 7 - Online table 5
- Data supplement 8 - Online table 6
- Data supplement 9 - Online table 7
Footnotes
-
Contributors All authors read and approved the final version of the manuscript. MMW: study concept and design, acquisition of data, analysis and interpretation of data, drafting of the manuscript. DL: technical and material support, critical revision of the manuscript for important intellectual content. MK: statistical analysis. IC: analysis and interpretation of data, critical revision of the manuscript for important intellectual content. LA, AD, PTS, JH, BO, PK, SVW, SM: acquisition of samples and characterisation of patients. PW, MS, RS: acquisition of samples and characterisation of patients, critical revision of the manuscript for important intellectual content. SV: critical revision of the manuscript for important intellectual content. GL: provided funding for validation phase, acquisition of samples. MD: acquisition of data, critical revision of the manuscript for important intellectual content. GD: obtained funding, acquisition of samples and data, critical revision of the manuscript for important intellectual content. GEB: study supervision, obtained funding, critical revision of the manuscript for important intellectual content.
-
Funding GlaxoSmithKline Pharmaceuticals has supported the data collection and the provision of samples and data from the UK/US/Canadian cohort. This work was financially supported by research grant G.0699.10N from the Fund for Scientific Research (FWO) Flanders, Belgium. Mira Wouters and Isabelle Cleynen are postdoctoral researchers and Séverine Vermeire is a senior clinical investigator of the FWO. Guy Boeckxstaens received research funding by a grant from the Flemish government (Odysseus Program, FWO). Research activities in Sweden were supported by funds from the Swedish Research Council to Mauro D'Amato. Magnus Simrén was supported by the Swedish Medical Research Council (grants 13409, 21691 and 21692) and The Marianne and Marcus Wallenberg Foundation. Magnus Simrén has received unrestricted research grants from Danone and AstraZeneca, and served as a Consultant/Advisory Board member for AstraZeneca, Danone, Novartis, Almirall and Shire/Movetis.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Ethical committee of GlaxoSmithKline, North Carolina, USA and the ethical committee of the Swedish hospitals.
-
Provenance and peer review Not commissioned; externally peer reviewed.