Article Text

Abstract
Objective Emerging evidence suggests that microRNA (miRNA)-mediated gene regulation influences a variety of autoimmune disease processes, including Crohn's disease (CD), but the biological function of miRNAs in CD remains unclear. We examine miRNA level in colon tissues and study the potential functions of miRNAs that regulate pathological genes during the inflammation process.
Design miRNA levels were assayed in the inflamed colon of 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced and IL-10 knockout (KO) chronic colitis mice and CD patients by microarray or qRT-PCR. The influence of differently expressed miR-141 on its putative target genes, CXCL12β, and leukocyte migration was investigated in colonic epithelia cells, colitis models and CD patients. The role of miR-141 was further studied in the experimental colitis mice by intracolonic administration of miR-141 precursors or inhibitors.
Results An inverse correlation between miR-141 and CXCL12β/total-CXCL12 was observed predominantly in the epithelial cells of the inflamed colons from colitic mice and CD patients. Further study demonstrated that miR-141 directly regulated CXCL12β expression and CXCL12β-mediated leukocyte migration. Upregulation or downregulation of miR-141 in the TNBS-induced or IL-10 KO colitic colon regulated leukocyte infiltration and alleviated or aggravated experimental colitis, respectively. Additionally, colonic overexpression of CXCL12β abolished the therapeutic effect of miR-141 in TNBS-induced colitis.
Conclusions This study showed that the pathway of miR-141 targeting CXCL12β is a possible mechanism underlying inflammatory cell trafficking during colonic inflammation process. Inhibiting colonic CXCL12β expression and blocking colonic immune cell recruitment by using miRNA precursors represents a promising approach that may be valuable for CD treatment.
- Chemokines
- Crohn'S Disease
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
-
microRNAs (miRNAs) Are differentially expressed in the colon tissue of Crohn's disease (CD) patients.
-
Increased expression of CXCL12, mainly by intestinal epithelial cells, has also been identified in the inflamed colons from inflammatory bowel disease patients and experimental colitis mice.
-
Elevated colonic CXCL12 is involved in intestinal inflammation via promoting immune cells infiltrations.
What are the new findings?
-
An inverse correlation between miR-141 and CXCL12β/total-CXCL12 is observed predominantly in the epithelial cells of the inflamed colons from CD patients and two different experimental colitic models (trinitrobenzene sulfonic acid induction and IL-10 null mice).
-
The upregulation of colonic total-CXCL12 is mainly contributed by CXCL12β during the intestinal inflammation.
-
miR-141 Regulates colonic leukocytic trafficking by targeting CXCL12β during intestinal inflammation.
-
Changing the colonic miR-141 level influences the process of experimental colitis and the therapeutic effect of miR-141 precursors is CXCL12β-dependent.
How might it impact on clinical practice in the foreseeable future?
-
This pathway of miR-141 targeting CXCL12β contributes significantly to the development of intestinal inflammation, likely by influencing the recruitment of immune cells into the inflamed colonic tissue. Inhibiting colonic CXCL12β expression and immune cell recruitment with miRNA represents a feasible and promising approach that may be valuable for the treatment of CD.
Introduction
Crohn's disease (CD), one of the two main subtypes of inflammatory bowel disease (IBD), is a chronic, relapsing and remitting inflammatory disorder within the gastrointestinal tract.1 Although the precise molecular basis of CD remains unclear, it is generally accepted that a combination of genetic susceptibility, environmental triggers, luminal bacterial flora disharmony and immune imbalance results in intestinal inflammation.2–4 Among these pathogenic factors, the dense infiltration of inflammatory cells into the inflamed colon plays an important role in the pathogenesis of CD.5 ,6 Increased chemokine expression in the colon tissue promotes the recruitment of immune cells to the inflamed site.7–9 Therefore, a significant amount of research has investigated the mechanisms that regulate chemoattractive protein expression in CD.10
MicroRNAs (miRNAs) are a group of endogenous, small, non-coding RNAs of approximately 22 nucleotides in length that regulate gene expression via binding to the 3′-untranslated region (3′-UTR) of target mRNA molecules at the post-transcriptional level.11 Interestingly, there is compelling evidence that a variety of miRNAs are involved in the regulation of immunity, such as the development and differentiation of immune cells and release of inflammatory mediators.12 miRNAs are differentially expressed in the serum and mucosa of IBD patients, suggesting that miRNA dysregulation may play an important role in the development of IBD.13–16 The putative target genes and biological functions of these differentially expressed miRNAs in IBD have not been extensively studied. Wu et al16 and Bian et al17 studied the pathways of miR-192 targeting MIP-2α and miR-150 targeting c-Myb, respectively. However, it is unknown whether restoring the balance of these aberrantly expressed miRNAs could influence the development of IBD, and there is no report on the use of miRNA precursors or inhibitors to successfully treat IBD or murine colitis.
In the present study, we measured the miRNA levels in the inflamed colons from 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced colitis mice, IL-10 knockout (KO) spontaneous colitis mice and CD patients. Differently expressed miR-141 in the inflamed colon of mouse and human was verified. The influence of miR-141 on its target CXCL12β, and inflammatory cell migration was investigated in colonic epithelia cells, two experimental colitis models and CD patients. The role of miR-141 was further studied in the experimental colitis models by intracolonic administration of miR-141 precursors or inhibitors.
Materials and methods
Human colon tissues, cell lines and chemical reagents
Colonoscopic pinch biopsies from normal sites and inflamed sites of the sigmoid colons of 15 CD patients (see online supplementary table S1) were obtained from Jiangsu Provincial People’s Hospital (Nanjing, China). Ethical approval was obtained from the local ethics committee. The CD diagnoses were confirmed by histopathological analysis on biopsies and a histopathological scoring system was used to evaluate the severity of CD patients.18 CT26 cells, Jurkat cells, THP-1 cells and RAW 264.7 cells (ATCC, Manassas, Virginia, USA) were cultured in Roswell Park Memorial Institute medium (RPMI)-1640 containing 10% fetal bovine serum (FBS) (Life Technologies, Grand Island, New York, USA). Caco-2 cells (ATCC) were cultured in Dulbecco's modified Eagle medium containing 10% FBS (Life Technologies). Mesenteric lymph node (MLN) cells, fresh bone marrow cells, primary mouse colonic epithelial cells (CECs), colonic immune cells and colonic smooth muscle cells (SMCs) were isolated and maintained as described in the online supplementary materials. Chemical reagents were purchased from Sigma (St Louis, Missouri, USA).
Establishment of experimental colitis
Female BALB/c mice, C3Bir.129P2 (B6)-Il10tm1Cgn/Lt (IL-10 KO) mice, C3H/HeJ mice were obtained from the Laboratory Animal Center of Nanjing University (Nanjing, China). All animals received care according to Chinese legal requirements.
TNBS-induced acute colitis in 6–8-week-old female BALB/c mice was carried out as reported previously.19 Briefly, 2 mg or 3 mg of TNBS (Sigma) in 100 μL 50% ethanol was slowly administered into the lumen of the colon via a catheter inserted 4 cm into the colon through the anus. Mice were then kept in a vertical position for 30 s. Control mice received 50% ethanol alone. Mice were sacrificed at day 3 after colitis induction.
Seven to eight weeks IL-10 KO mice were maintained under conventional (non-pathogen-free) conditions for 12 weeks. C3H/HeJ mice were used as controls. At 20 weeks, IL-10 KO mice and control mice were sacrificed. Serum amyloid A (SAA) was used to evaluate the severity of colitis.
The behaviour of the mice in both the models was observed daily, and the colons were excised for macroscopic observation, histopathological analysis, qRT-PCR assay, western blot analysis and chemokines/cytokines analysis. The disease activity index (DAI) was used to evaluate the grade of intestinal inflammation in TNBS-induced colitis.20 For histopathological examination, histological score of H&E staining sections was graded from 0 to 4 according to a previous report.20
In vitro validation and chemotaxis assays
For the target validation of miR-141, cell transfection/infection assay and luciferase reporter assay were performed. For functional study, in vitro chemotaxis assays were performed. MLN cells, fresh bone marrow cells, RAW 264.7 cells, Jurkat cells and THP-1 cells (2×105/100 μL of RPMI 1640 containing 0.1% bovine serum albumin) were respectively transferred to the upper layer of a 5 μm-pore polycarbonate membrane (Transwell; Corning, Corning, New York, USA). The bottom chambers contained 0.6 mL of media that was collected from the supernatants of 1×106 primary CECs from control mice or TNBS mice or Caco-2 cells 24 h after pre-miR-141/anti-miR-141 transfection. After 5 h of incubation, migrating cells were collected from the lower wells and counted by flow cytometry using antimouse-Gr-1 fluorescein isothiocyanate (FITC), antimouse-F4/80 FITC, antimouse-CD3 FITC, antihuman-CD3 FITC and antihuman-CD18 FITC (eBioscience, San Diego, California, USA). The number of cells in the lower wells out of the total number of cells in the upper well yielded the percentage of cell migration. The migration index was the percentage of cell migration to the lower well containing the medium from CECs or Caco-2 cells with different treatments compared with the migration to the lower well containing the medium from control CECs or Caco-2 cells with mock transfection. In some experiments, recombinant CXCL12β was added to the lower chamber containing supernatant from CECs or Caco-2 cells that underwent pre-miR-141 pretreatment, or antibody against CXCL12 was added to the lower chamber containing supernatant from CECs or Caco-2 cells that underwent anti-miR-141 pretreatment. In preliminary experiments, the promotion or inhibition of immune cells toward the lower chamber was achieved using 5 ng/mL mouse or human CXCL12β recombinant protein (Abnova, Taipei, China) or 1 μg/mL anti-CXCL12 (R&D Systems, Minneapolis, Minnesota, USA), respectively; thus, these concentrations were selected for the promotion or inhibition studies (data not shown).
Microarray and qRT-PCR assay
The microarray and qRT-PCR assay are described in the online supplementary material. The primer sequences for qRT-PCR of miRNAs and genes are shown in online supplementary table S2.
Treatment and assessment of colonic inflammatory changes
Detailed protocols describing the treatment and examination of experimental colitis model are shown in the online supplementary material.
Statistics
The results are expressed as the means±SEMs. Hierarchical cluster analysis was conducted using Gene Cluster software (Stanford University). Significance analysis of microarray (SAM) was conducted. To exclude extreme outliers, miRNAs with expression levels lower than a threshold (CT miRNA—CT U6>15, mean fold change >1.5 or <0.67) were eliminated. The differences between groups were analysed using a Mann–Whitney U test and, if appropriate, by a Kruskal–Wallis analysis of variance test. A value of p<0.05 was considered significant.
Results
Validation of differentially expressed miRNAs in the inflamed colon of mouse and human
We established TNBS-induced and IL-10 KO colitis models to determine whether miRNAs are differentially expressed in colitis. The pathological features of TNBS colitis (colon hyperaemia and transmural inflammation) and IL-10 KO mice (diarrhoea and massive colonic leukocyte aggregation) agree with previous studies (figure 1A).

microRNA (miRNA) profiling analysis in trinitrobenzene sulfonic acid (TNBS)-induced and IL-10 knockout (KO) colitis. (A) Success in establishing TNBS colitis resulted in colon hyperaemia and transmural inflammation. IL-10 KO mice exhibited diarrhoea and massive colonic leukocyte aggregation. Scale bar, 100 μm. (B) Cluster analysis of miRNA induced or repressed (q<0.056, fold>1.5) in TNBS and IL-10 KO colitis. (C) The relative miRNA expression ratios of experimental colitis to control in the qRT-PCR data compared with microarray data. n=5 mice per group. (D) qRT-PCR analysis of miR-31, miR-2137 and miR-141 in colon biopsy tissues from Crohn's disease (CD) patients between normal sites and inflamed sites; n=15 CD patients. Values are expressed as the means (SEMs). *p<0.05. NS, no significant change.
The miRNA levels in the inflamed colon of TNBS-induced and IL-10 KO mice were screened by microarray. SAM analysis generated 21 differentially expressed miRNAs in TNBS colitis (see online supplementary table S3) and 35 differentially expressed miRNAs in IL-10 KO mice (see online supplementary table S4). Hierarchical cluster analysis of the changed miRNAs identified by SAM generated a dendrogram with two major branches in both columns (colitis vs control) and rows (upregulated vs downregulated miRNAs) in both experimental colitis models (figure 1B). We further performed qRT-PCR to quantify 10 differentially expressed miRNAs in another five pairs of colitis and control samples. Consistent with the microarray data, the qRT-PCR analysis revealed the same trends in expression for these miRNAs in both experimental models (figure 1C). The intersection analysis of miRNA profiling demonstrated that miR-31, miR-2137 and miR-141 were similarly dysregulated in both experimental models. Clinical data demonstrated that the change of miR-31 and miR-141 in the inflamed colon of CD patients had the same change trend as those in the experimental colitis models (figure 1D).
Correlationship analysis between miR-141 and CXCL12β
To study the possible mechanisms of dysregulated miRNA underlying intestinal inflammation, three algorithms, TargetScan, miRanda and PicTar, were combined to search for target genes of miR-141 and miR-31. A list of miRNA-target genes potentially involved in colitis was generated (see online supplementary table S5). When studying CXCL12, STAT5a, GATA6, PTEN, MAP2K4 and PDCD4 as target genes of miR-141, we found that the levels of STAT5a, GATA6, PTEN and MAP2K4 had no converse relationship with miR-141 in the experimental colitis models (see online supplementary figure S1A). PDCD4 has also been proved not a target of miR-141.21 Whereas, strong evidences suggest that the upregulated CXCL12 promotes the leukocytic infiltration during the mucosal inflammation and blocking the CXCL12/CXCR4 axis ameliorates colonic inflammation in experimental colitic model, indicating that CXCL12 as target of miR-141 may regulate mucosal inflammation in CD.22 ,23
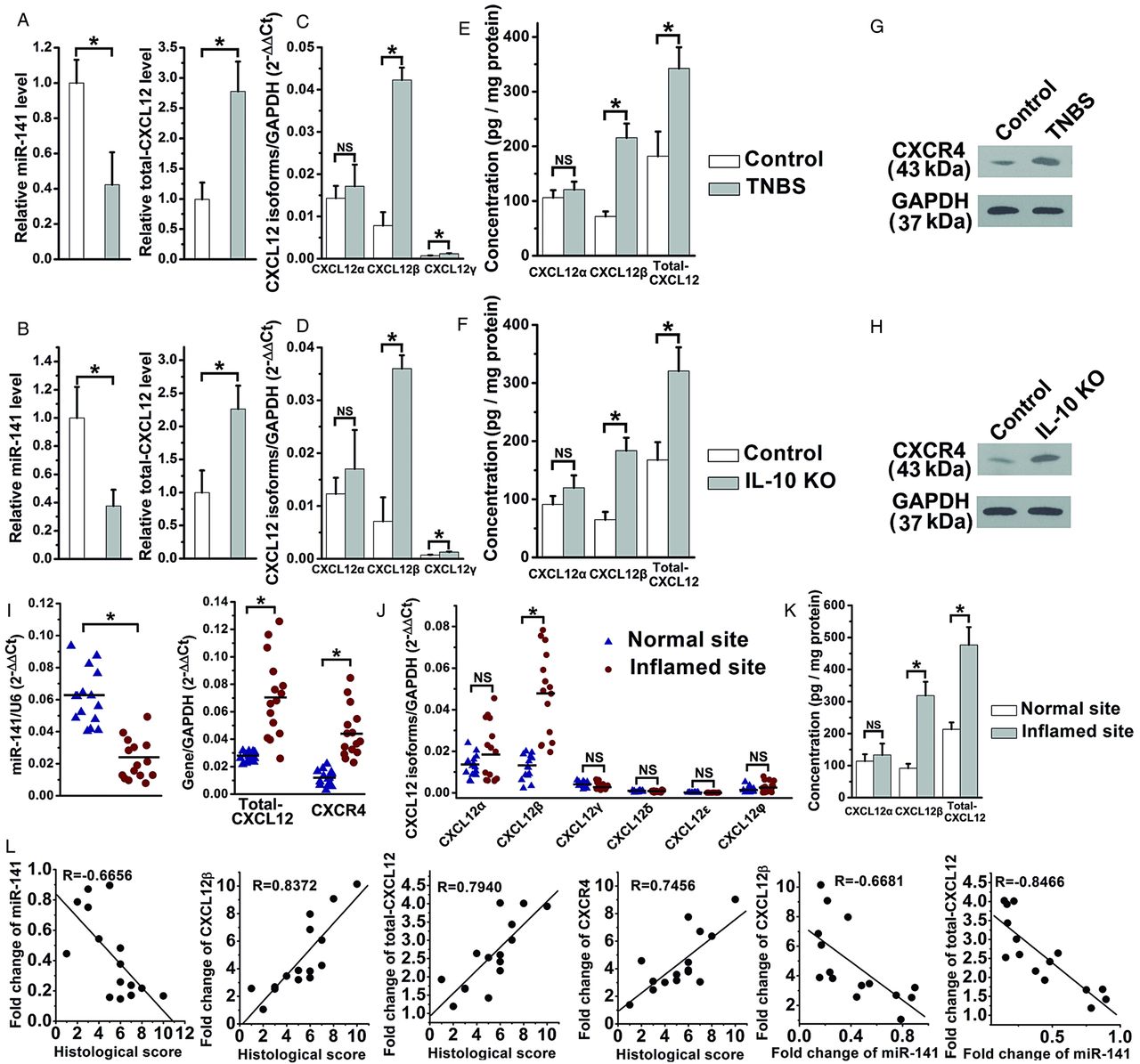
Three mouse CXCL12 isoforms (α, β and γ) and six human CXCL12 isoforms (α, β, γ, δ, ɛ and φ) have been identified; whereas only mouse CXCL12β/γ and human CXCL12β were predicted to be the target of miR-141.24 ,25 Hybrids between the mice CXCL12β/γ or human CXCL12β 3′UTR and miR-141 are shown in online supplementary figure S2A. The levels of total-CXCL12, CXCL12β/γ and CXCR4 were significantly increased and miR-141 level was reduced in the inflamed colon of TNBS-induced and IL-10 KO mice (figure 2A–H). CXCL12β was observed more abundant than CXCL12γ in murine colon (figure 2C,D), therefore, the elevated level of colonic total-CXCL12 was mainly contributed by the upregulation of CXCL12β in the murine colitis models. The qRT-PCR assay demonstrated a decreased level of miR-141 and increased mRNA levels of CXCL12β, total-CXCL12 and CXCR4 in the inflamed sites of 15 CD patients (figure 2I,J). Significantly increased protein levels of CXCL12β, total-CXCL12 rather than CXCL12α were also observed in the inflamed colon of 15 CD patients (figure 2K). CXCR7 showed no significant change during the colonic inflammation in both mouse and human (data not shown). Via the Pearson's correlation analysis, there was an inverse relationship between miR-141 and histopathological score, and a positive relationship between CXCL12β rather than other isoforms, total CXCL12, CXCR4 and histopathological score in CD patients. The inverse correlation between miR-141 and CXCL12β/total-CXCL12 was also illustrated using Pearson's correlation scatter plots (figure 2L, see online supplementary figure S2B,C).
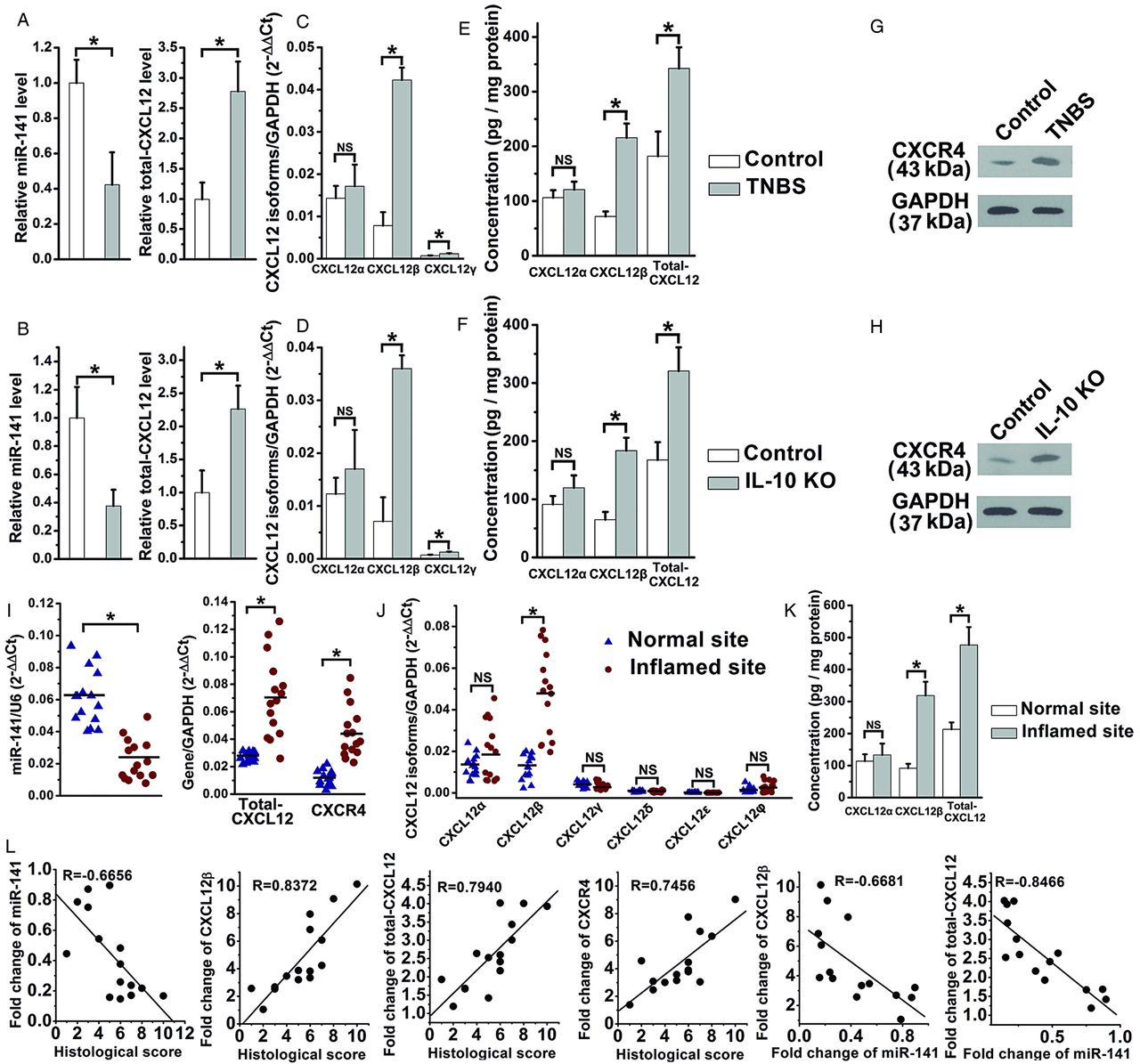
The inverse correlation between CXCL12β/total-CXCL12 and miR-141 in experimental colitis and Crohn's disease (CD) patients. qRT-PCR analysis of the expression levels of miR-141/total-CXCL12 (A and B), different CXCL12 isoforms (C and D), ELISA analysis of the protein levels of CXCL12α/CXCL12β/total-CXCL12 (E and F), western blotting analysis of CXCR4 (G and H) in trinitrobenzene sulfonic acid (TNBS)-induced and IL-10 knockout (KO) colitis; n=9-11 mice per group. The mRNA levels of miR-141, total-CXCL12, CXCR4 (I), CXCL12 isoforms (J) and the protein levels of CXCL12α/CXCL12β/total-CXCL12 (K) in 15 CD patients. (L) Pearson's correlation scatter plot of the fold changes of miR-141, CXCL12β, total-CXCL12, CXCR4 mRNA with histological score, and the fold changes of miR-141 with CXCL12β, total-CXCL12 in CD patients. Values are expressed as the means (SEMs). *p<0.05.
Besides miR-141 target CXCL12β, we have examined miRNAs possibly involved in the regulation of CXCL12 and CXCR4 in the murine colitis model.26–32 As shown in online supplementary figure S1B, none of miR-126-3p, miR-127, miR-140, miR-23a, miR-23b, miR-222, miR-223 and miR-27a show a converse correlation with CXCL12β/total-CXCL12. As shown in online supplementary figure S1C, miR-146a or miR-139-5p may contribute, respectively, to the upregulation of CXCR4 in TNBS colitic mice or in IL-10 KO mice.
Cellular localisation of miR-141 and CXCL12β in mouse and human colon
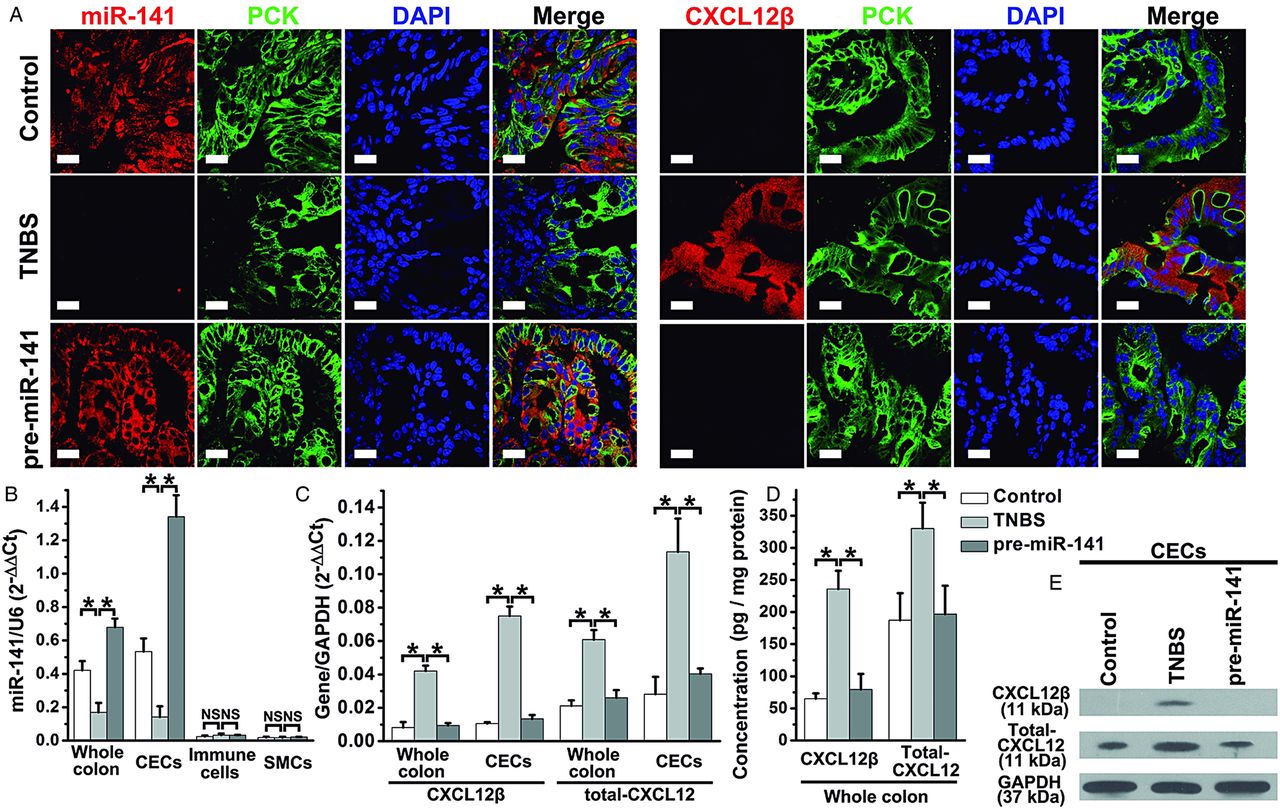
To identify the cell types expressing miR-141 and CXCL12β, we performed in situ hybridisation/immunofluoresence staining on colon sections; miR-141 was predominantly expressed in epithelial cells (pan-cytokeratin+ (PCK+) cells) of the normal colon of mouse and human (figure 3A) but not in immune cells or SMCs (see online supplementary figure S3). miR-141 In epithelial cells was significantly decreased in the murine colitis colon and the inflamed colon of CD patients (figure 3A). Additionally, CXCL12β protein was mostly localised in epithelial cells in TNBS-induced colitis colon and the inflamed sites of CD patients but was detected at very low level in epithelial cells in healthy colon tissues, suggesting that CXCL12β and miR-141 are predominantly expressed in the same cell types in the mouse/human colon but in opposing states (figure 3A). The results of qRT-PCR and western blotting analysis on different cell types from colon also indicated a negative correlation between miR-141 and CXCL12β/total-CXCL12, which was observed in CECs rather than colonic immune cells or SMCs (figure 3B–E).

miR-141 and CXCL12β localisation in trinitrobenzene sulfonic acid (TNBS) colitic colon and human colon tissues. (A) Immunofluorescent staining with different cell markers and in situ hybridisation for miR-141 were performed on colon sections from TNBS colitis and Crohn's disease (CD) patients (red, miR-141; green, pan-cytokeratin (PCK); blue, 4′,6-diamidino-2-phenylindole (DAPI) nuclear staining). Dual-immunofluorescence staining with different cell markers and CXCL12β were used to investigate CXCL12β localisation (red, CXCL12β; green, PCK; blue, DAPI nuclear staining). Scale bar, 20 μm. The levels of miR-141 (B), CXCL12β mRNA (C), total-CXCL12 mRNA (D), CXCL12β/total-CXCL12 protein levels (E) in different types of cells isolated from a TNBS colitic colon. Values are expressed as the means (SEMs). *p<0.05. NS: no significant change; n=8–10 mice per group.
In vitro validation of CXCL12β as a target of miR-141
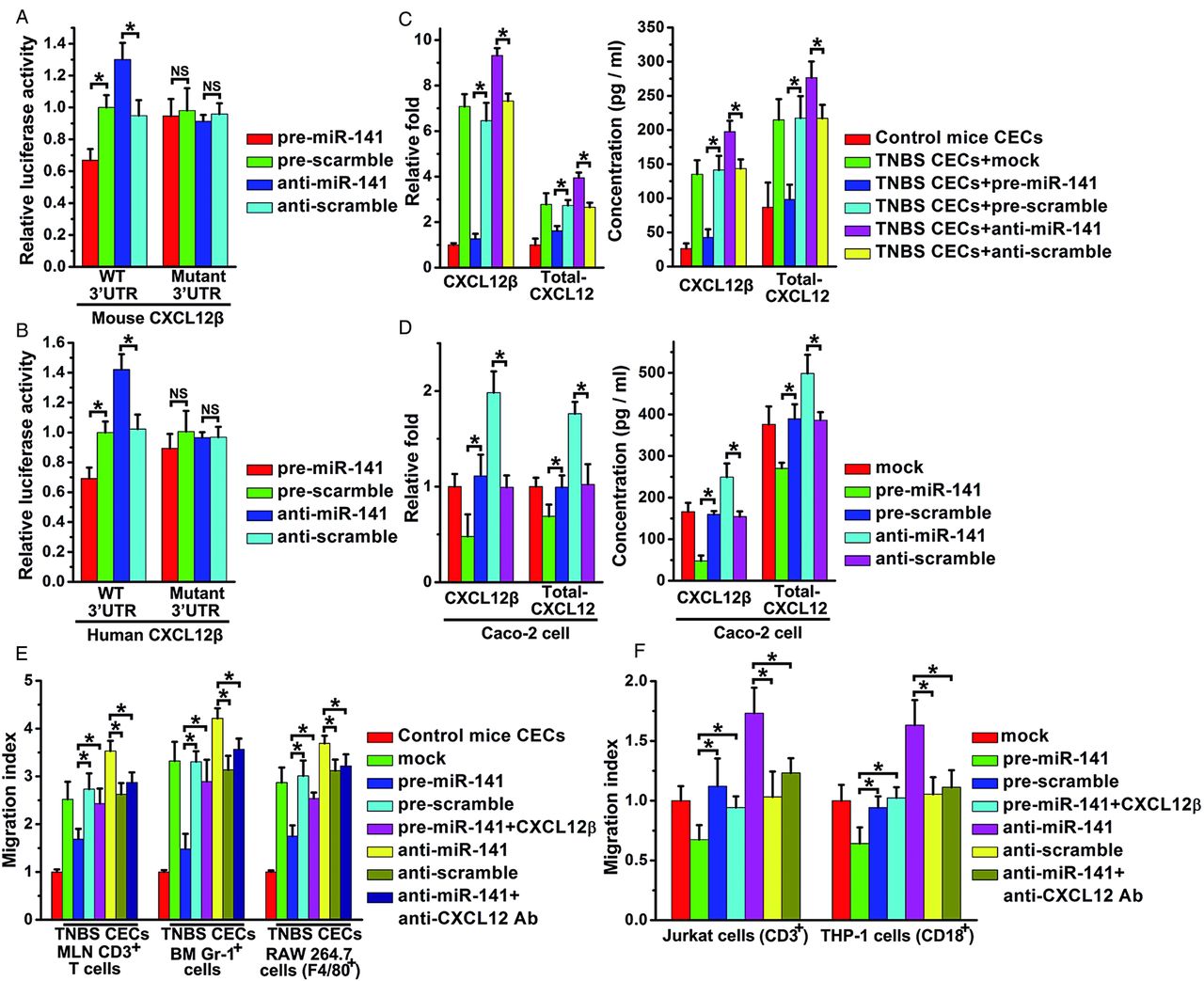
Overexpression of miR-141 suppressed luciferase reporter activity that contains the full-length 3′UTR of the mouse CXCL12β/γ or human CXCL12β (figure 4A,B, see online supplementary figure S4A), but had no effect on luciferase reporter activity that contains other isoforms (see online supplementary figure S4A,B). Mutagenesis of the seed sequences of the two predicted miR-141 binding sites restored luciferase expression, thereby confirming the specificity of the interaction between miR-141 and the mouse CXCL12β/γ or human CXCL12β 3′UTR. Meanwhile, compared with the control groups (figure 4A,B, see online supplementary figure S4A), inhibition of miR-141 resulted in an increase in reporter activity of mouse CXCL12β/γ or human CXCL12β.
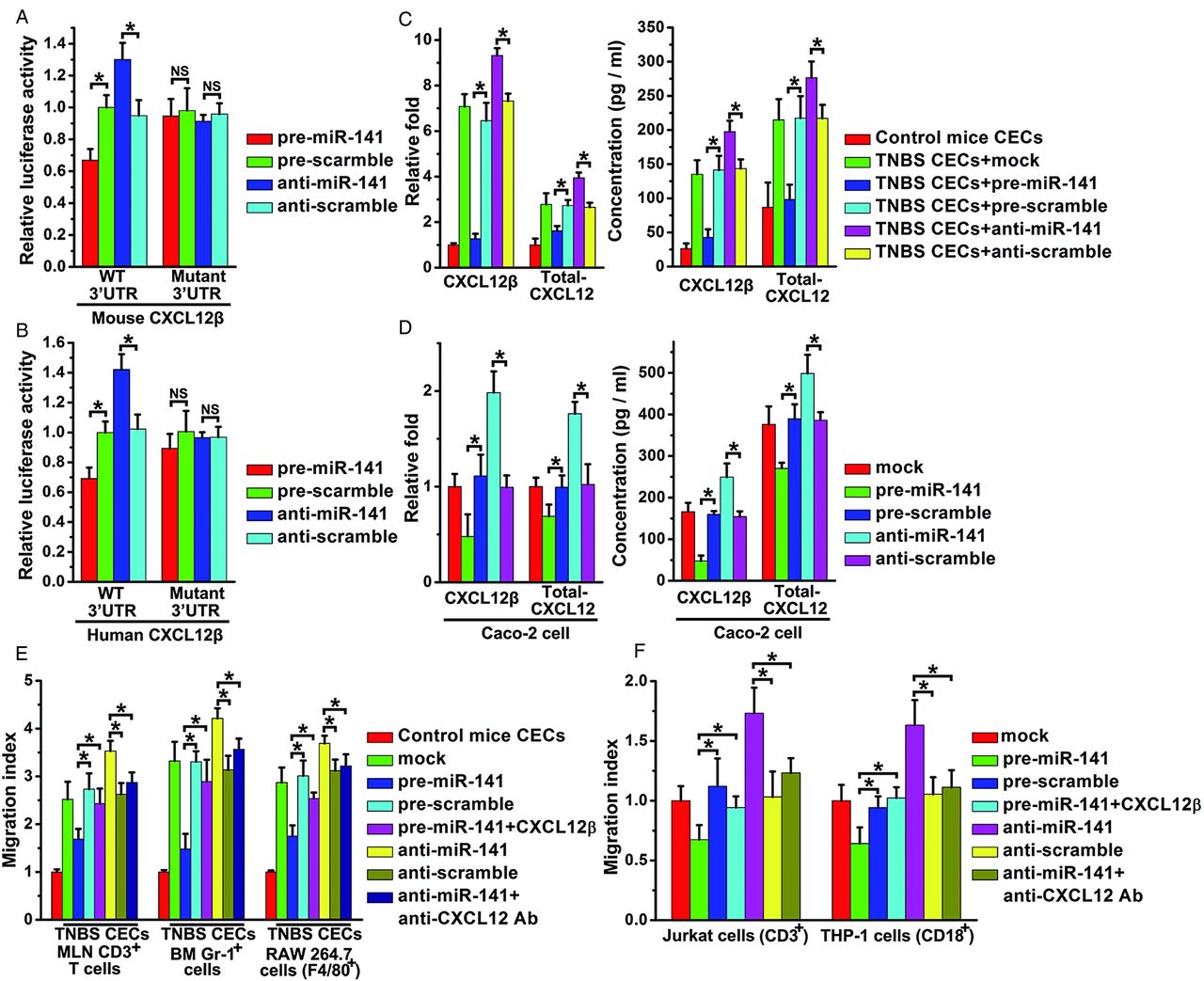
Identification of CXCL12β as a target of miR-141 and migration assay. The relative luciferase activity containing full length 3′untranslated region (3′UTR) of mouse/human CXCL12β in primary mouse colonic epithelial cells (CECs) (A) or Caco-2 cells (B) was assayed after transfection with pre/anti-miR-141 or scrambled pre/anti-miR-141. The binding site mutant plasmid (Mutant) were also used. The relative CXCL12β/total-CXCL12 mRNA level and CXCL12β/total-CXCL12 concentrations in the supernatants from normal mouse CECs and trinitrobenzene sulfonic acid (TNBS) mouse CECs (C) or Caco-2 cells (D) after mock, pre/anti-miR-141 or scrambled pre/anti-miR-141 transfection. Mesenteric lymph node T cells (MLN CD3 cells), bone marrow granulocytes (Gr-1+ cells), mouse monocytes (RAW 264.7 cells) (E) or human T cells (Jurkat cells), human monocytes (THP-1 cells) (F) toward supernatants of CECs from control mice or TNBS mice or Caco-2 cells with mock, pre/anti-miR-141 or scrambled pre/anti-miR-141 transfection. In some experiments, CXCL12β recombinant protein or anti-CXCL12 antibody (Ab) was added to the upper well. Migration index: percentage of migration of cells to the lower well compared with migration to the medium from control CECs or Caco-2 cell with mock transfection. Values are expressed as the means (SEMs). Five samples were analysed per condition, and experiments were performed in triplicate. *p<0.05.
The correlation between miR-141 and CXCL12β/total-CXCL12 was further examined in mouse CECs or human colon epithelial Caco-2 cells which normally express CXCL12.33 ,34 Upregulation of miR-141 significantly reduced mRNA levels of CXCL12β/CXCL12γ/total-CXCL12 in mouse CECs (figure 4C) or CXCL12β/total-CXCL12 in Caco-2 cells (figure 4D) and the protein levels of supernatant CXCL12β/total-CXCL12 in CECs or Caco-2 cells (figure 4C,D), but had no effect on that of other isoforms (data not shown). However, miR-141 inhibitors upregulated CXCL12β/total-CXCL12 transcript and protein levels in TNBS-induced CECs or Caco-2 cells (figure 4C,D).
We also performed target validation in CT26 cells cotreated with lentivirus and pre/anti-miR-141. Both miR-141 mimics and inhibitors changed the miR-141 level in CT26 cells (see online supplementary figure S4C). As shown in online supplementary figure S4D,E, pre-miR-141 and anti-miR-141, respectively, downregulated or upregulated the level of CXCL12β in CT26 cells that were preinfected with CXCL12β lentivirus rather than in CT26 cells treated with CXCL12β 3′UTR mutant lentivirus. The viability of primary CECs, Caco-2 and CT26 cells with different treatments was examined using a CCK-8 kit, and the viability was higher than 90%.
miR-141 Regulates leukocyte migration toward the CEC-derived supernatants
Leukocyte chemotaxis assay was performed to investigate the effect of miR-141 targeting CXCL12β on leukocyte migration. Supernatants were collected from the normal CECs, TNBS CECs or Caco-2 cells that were transfected with pre/anti-miR-141. Migration analysis demonstrated that the chemotactic responses of mouse mesenteric CD3 T cells, Gr-1+ cells (mouse bone marrow granulocytes), RAW 264.7 cells (mouse monocytes), Jurkat cells (human T cells) and THP-1 cells (human monocytes) were significantly suppressed by pre-miR-141, whereas anti-miR-141 treatment enhanced the migration index of these immune cells. By contrast, CXCL12β recombinant protein added to the supernatant of pre-miR-141 treated CECs or Caco-2 cells recovered the migration rate that was reduced by pre-miR-141 treatment, (figure 4E,F) whereas, anti-CXCL12 antibody added to the supernatant of anti-miR-141 treated CECs or Caco-2 cells reduced the migration rate that was upregulated by anti-miR-141 treatment (figure 4E,F).
Cellular localisation of miR-141 after pre-miR-141 administration
Frozen sections prepared from the colons of Cy3 miRNA precursor treated mice were stained with epithelial cell markers. As shown in online supplementary figure S5, most of Cy3 miRNA precursors accumulated in the CECs (PCK+ cells). In situ hybridisation and immunofluorescent staining were also demonstrated that the intracolonic administration of pre-miR-141 significantly increased the miR-141 level and inhibited CXCL12β expression in CECs of TNBS-colitic colon (figure 5A). To further quantify the cellular level of miR-141, CECs, colonic immune cells and SMCs were isolated at 12 h after pre-miR-141 injection for qRT-PCR and western blotting analysis. Intracolonic administration of pre-miR-141 significantly upregulated miR-141 level (figure 5B) and downregulated CXCL12β/total-CXCL12 in whole colon and CECs (figure 5C–E).

Cellular levels of miR-141, CXCL12β and total-CXCL12 after pre-miR-141 administration. (A) Immunofluorescent staining with colonic epithelial cell (CEC) marker (pan-cytokeratin (PCK)) and in situ hybridisation for miR-141 were performed on colon sections from trinitrobenzene sulfonic acid (TNBS) colitis after pre-miR-141 administration (red, miR-141; green, PCK; blue, DPAI nuclear staining). Dual-immunofluorescence staining with CEC marker (PCK) and CXCL12β for CXCL12β localisation were performed on TNBS colitic colon sections after pre-miR-141 administration (red, CXCL12β; green, PCK; blue, DPAI nuclear staining). Scale bar, 20 μm. The levels of miR-141 (B), CXCL12β/total-CXCL12 mRNA (C) and CXCL12β/total-CXCL12 protein level in whole colon (D) or CECs isolated from TNBS colitic colon (E) after pre-miR-141 administration. The values are expressed as the means±SEM; n=5–6 mice per group. *p<0.05. NS, no significant change.
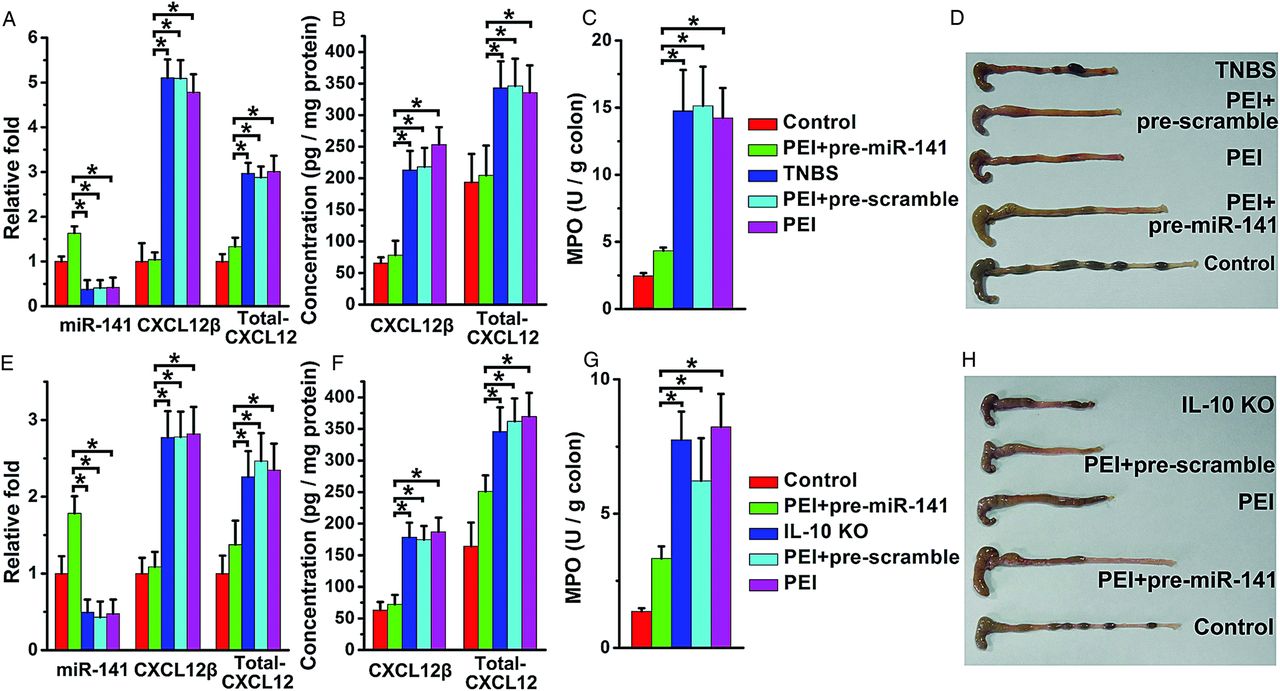
Pre-miR-141 treatment protects against the development of TNBS-induced and IL-10 KO colitis
As shown in figure 6A,B, pre-miR-141 intracolonic administration significantly increased miR-141 levels and suppressed CXCL12β/total-CXCL12 expression in TNBS-colitic colon. Mice treated with pre-miR-141 rapidly recovered the lost body weight induced by TNBS and experienced lower DAI levels (see online supplementary figure S6A,B). After pre-miR-141 treatment, colonic myeloperoxidase (MPO) activity, correlating with mucosal neutrophils infiltration, was significantly reduced, and the colon showed no significant signs of macroscopic inflammation (figure 6C,D). Histological examination showed that TNBS-induced colitis affected all layers of the colon, including submucosal oedema and strong leukocyte infiltration. After mice were treated with pre-miR-141, a striking improvement of histological signs became apparent (see online supplementary figure S6C,E).
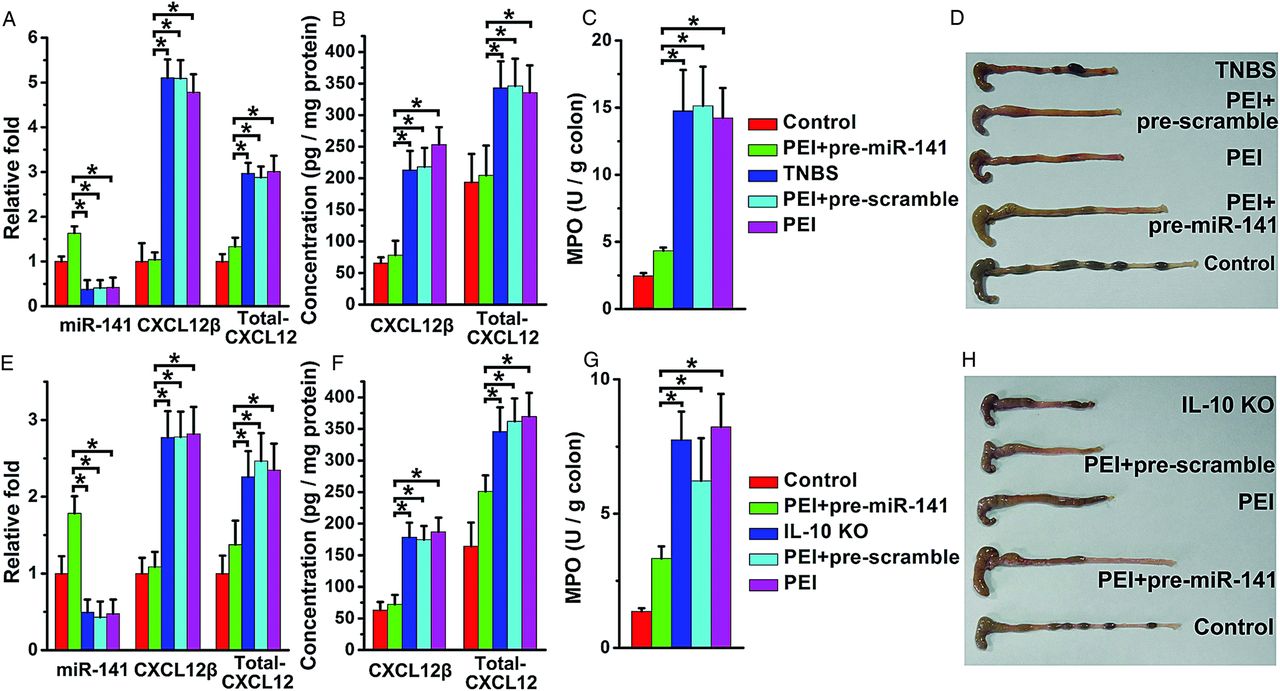
pre-miR-141 treatment ameliorated trinitrobenzene sulfonic acid (TNBS)-induced and IL-10 knockout (KO) colitis. The levels of miR-141, CXCL12β/total-CXCL12 mRNA level (A), CXCL12β/total-CXCL12 protein level (B) in the colons from TNBS colitic mice with pre-miR-141 treatment. Clinical evolution and severity were monitored by myeloperoxidase (MPO) activity determination (C) and colon pictures (D). The levels of miR-141, CXCL12β/total-CXCL12 mRNA level (E), CXCL12β/total-CXCL12 protein level (F) in the colons from IL-10 KO mice with pre-miR-141 treatment. The therapeutic effect of pre-miR-141 in IL-10 KO mice was observed through MPO activity determination (G) and colon photographs (H); n=8–11 mice per group. Values are expressed as the means (SEMs). *p<0.05.
IL-10 KO mice developed spontaneous colitis with apparent diarrhoea and increased SAA. After repeated treatment with miR-141, colonic miR-141 levels were upregulated, and CXCL12β/total-CXCL12 expression was downregulated (figure 6E,F). Colonic MPO activity, macroscopic observation (figure 6G,H), SAA and histologic examination (see online supplementary figure S7A,B,D) demonstrated that pre-miR-141-treated IL-10 KO mice developed less serve colitis. The favourable therapeutic effect of miR-141 in the two experimental models was associated with diminished inflammatory cytokines production and reduced leukocyte (T cells, neutrophils and macrophages) infiltration (see online supplementary figures S6D,E and 7C,D).
Anti-miR-141 aggravates TNBS-induced colitis
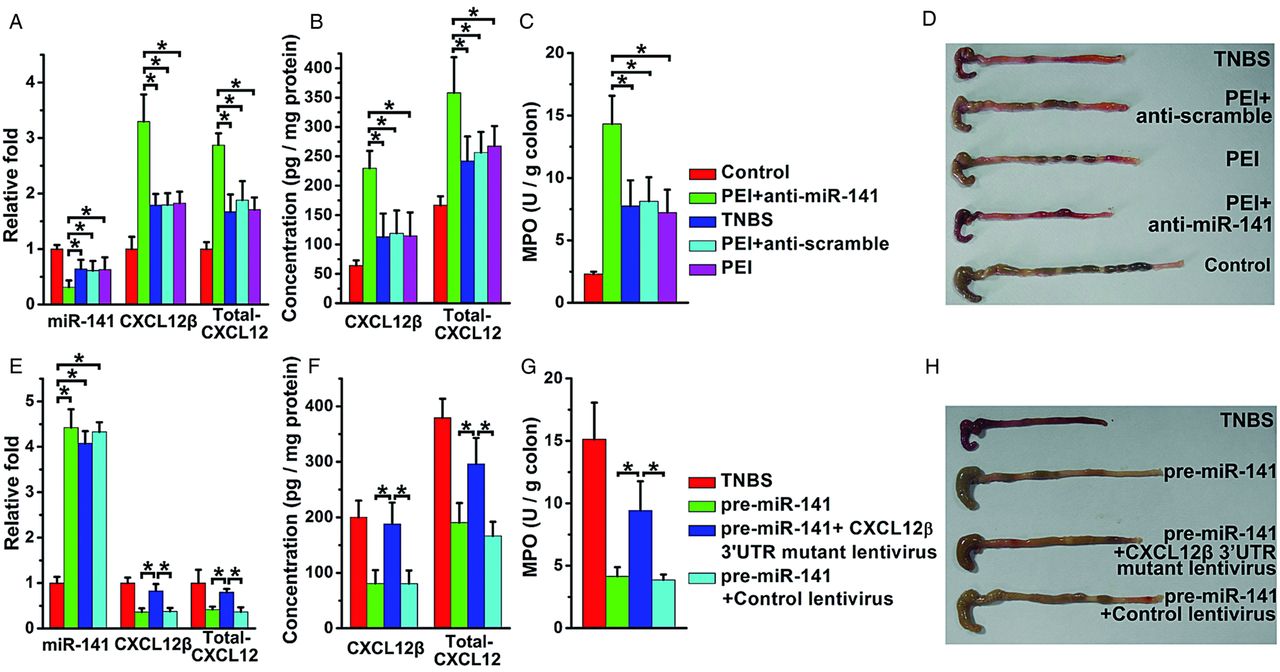
To confirm the role of miR-141 in TNBS-induced colitis, we next administered anti-miR-141 to mice that had been given a low dose of TNBS to induce moderate colitis. Anti-miR-141 further reduced the colonic miR-141 level and increased CXCL12β/total-CXCL12 expression (figure 7A,B). MPO activity, colon pictures, body weight changes, DAI and histologic examinations of the colons showed that mice given anti-miR-141 developed more severe colitis (figure 7C,D, see online supplementary figure S8A–C,E), which was associated with enhanced expression of proinflammatory cytokines and diminished immune cell expansion (see online supplementary figure S8D).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mice injected with anti-miR-141 develop severe trinitrobenzene sulfonic acid (TNBS)-induced colitis and colonic overexpression of CXCL12β prevented the therapeutic effect of pre-miR-141 in TNBS colitic mice. The levels of miR-141, CXCL12β/total-CXCL12 mRNA level (A), CXCL12β/total-CXCL12 protein level (B) in the colons from TNBS colitic mice with anti-miR-141 treatment. Clinical evolution and severity were monitored by myeloperoxidase (MPO) activity determination (C) and colon pictures (D). The levels of miR-141, CXCL12β/total-CXCL12 mRNA level (E), CXCL12β/total-CXCL12 protein level (F) of TNBS colitic mice treated with CXCL12β 3′untranslated region (3′UTR) mutant lentivirus and pre-miR-141. Clinical evolution and severity were monitored daily by MPO activity determination (G) and colon pictures (H); n=8–11 mice per group. Values are expressed as the means (SEMs). *p<0.05.
The therapeutic effect of pre-miR-141 on TNBS-induced colitis is mediated by CXCL12β suppression
Because administration of pre-miR-141 to colitic mice leads to the downregulation of CXCL12β, a chemokine that is supposed to exert pathogenic effects in colitis, we examined whether pre-miR-141 alleviated experimental colitis through the inhibition of CXCL12β. A CXCL12β 3′UTR-mutant lentivirus was intracolonically administered prior to the induction of TNBS. The lentivirus significantly improved colonic CXCL12β/total-CXCL12 expression when compared with TNBS mice with control lentivirus treatment or without any treatment (figure 7E,F). Due to its 3′UTR binding site mutant, the overexpression of CXCL12β by this lentivirus vector cannot be regulated by miR-141 (see online supplementary figure S4H,I). The overexpression of colonic CXCL12β with this lentivirus vector abolished the anti-inflammatory effect of pre-miR-141 on TNBS-induced colitis (figure 7G,H, see online supplementary figure S9A–C,E). Moreover, the CXCL12 3′UTR-mutant lentivirus treatment significantly reduced the inhibitor effect of pre-miR-141 on colonic cytokine secretion and leukocyte infiltration (see online supplementary figure S9E).
Discussion
In the present study, we focused on understanding the importance of miR-141 for the control of colonic inflammation; miR-141, a member of the miR-200 family, has been associated with a variety of human malignancies, such as prostate cancer and gastric cancer.35 ,36 Another study demonstrated that miR-141 is involved in epithelial-to-mesenchymal transition (EMT), which implicates its role in embryonic development and fibrosis.37 ,38 Additionally, miR-141 is aberrantly expressed in two other autoimmune diseases (lupus and psoriasis).39 ,40 However, the current studies on miR-141 has been restricted to tumorigenesis and EMT, and its immune function, especially in intestinal inflammation, remains unclear. A previous study detected a slight but not significant reduction of miR-141 in active ulcerative colitis patients.16 We also demonstrated that miR-141 was primarily expressed in CECs of the normal colon, and a significant reduction of miR-141 was observed in CECs from the TNBS colitic mice and the inflamed sites of CD patients. Although the reason for the reduced miR-141 in inflamed colon remains unclear, there is evidence that TNF-α stimulation can induce a significant reduction of miR-141 in human intestinal epithelial HT29 cells.16 We speculate that proinflammatory cytokine stimulation during the inflammation process contributes to the downregulation of miR-141 in CECs. Its regulation by TNF-α, and its aberrant expression in CD patients and experimental colitis imply that miR-141 plays a critical role in intestinal inflammation.
CXCL12β, an epithelial cell-expressed chemokine, has been verified a target of miR-141 and can be upregulated due to a reduction of miR-141 in the inflammation process in our study. Compared with the ubiquitously expressed splicing variant CXCL12α, CXCL12β has more resistance to proteolysis in the blood due to its four additional residues in the C-terminus.41 Therefore, CXCL12β can maintain a potent concentration gradient in the blood, which is more efficient to recruit circulating leukocytes than CXCL12α.24 Previous study indicates that CXCL12, mainly expressed by CECs, has been identified in the inflamed colons from IBD patients.23 We further demonstrate that the upregulation of colonic total-CXCL12 expression are mostly contributed by increased CXCL12β from CECs during mucosal inflammation. The significant chemotactic effect of CXCL12 is supported by reports demonstrating that IBD CEC-derived supernatants promote peripheral blood T cell and lamina propria T cell migration, and this effect can be abolished by specific antibodies against CXCL12 or its receptor CXCR4.23 CXCR4 level in the inflamed colon has also been observed to be increased in previous findings and our study.42 Therefore, during the process of intestinal inflammation, immune cells, including leukocytes, monocytes and neutrophils, which are attracted by the highly expressed CXCL12, infiltrate into the inflamed colon and secrete large volumes of proinflammatory cytokines and other types of chemokines. These pathological chemokines secreted by infiltrating inflammatory cells recruit more effector cells, which amplify the mucosal inflammation and lead to an exacerbation of the destructive processes within the colon. Thus, the CXCL12 secretion by CECs, leading to a chemokine–inflammatory cell self-reinforcing feedback loop, is one of the most important pathogenic mechanisms involved in the initiation and exacerbation of mucosal inflammation in IBD. Blocking this positive-feedback loop through selective manipulation of the CXCL12/CXCR4 axis will be valuable as a potential therapeutic strategy in IBD. TF14106, a CXCR4 antagonist, protects against DSS-induced colitis and colonic inflammation in IL-10 KO mice by inhibiting leukocyte recruitment into the inflamed intestine by blocking the colonic CXCL12/CXCR4 axis.22 Additionally, CXCL12 enhances CD4 T cell survival by inhibiting its spontaneous apoptosis, which also contributes to the perpetuation of intestinal inflammation.43 All this evidence demonstrates that CXCL12 is an important pathogenic factor involved in the mucosal inflammation process, and identifying its potential regulating mechanisms may provide promising targets for new drug development in the treatment of CD.
It has been reported that various factors, such as Wnt signalling, cell-contact conjunctions, DNA methylation and miRNAs, contribute to the regulation of CXCL12; however, the mechanism that promotes colonic CXCL12 expression during the process of intestinal inflammation remains unclear.44–46 Based on the fact that the level of CXCL12β is significantly increased in the inflamed CECs, and that miR-141 is also significantly reduced in the same cell type, we hypothesise that the miR-141 targeting CXCL12β pathway in CECs serves as an important mechanism to regulate colonic CXCL12β/total-CXCL12 expression and control immune cell trafficking during both experimental colitis models and the CD dysfunction process in humans. In vitro chemotaxis assays indicated that the inhibitory effects of pre-miR-141 and promoting effects of anti-miR-141 on the migration of immune cells toward human Caco2 cells or TNBS-stimulated mouse CEC-derived supernatants could be abrogated by the addition of recombinant CXCL12β or anti-CXCL12 antibody. In vivo, polyetherimide, a widely used gene carrier, effectively transferred pre-miR-141/anti-miR-141 into CECs, which are the main source of colonic CXCL12β, regulated colonic CXCL12β protein levels and attenuated or enhanced the CXCL12β-mediated chemokine-inflammatory cell-chemokine self-promoted feedback loop during the initial phase of inflammation, resulting in an amelioration or aggravation of experimental colitis models. Additionally, colonic CXCL12β overexpression abolished the therapeutic effect of pre-miR-141 in TNBS-induced colitis. These results prove that miR-141-mediated immune cell trafficking through CXCL12β in CECs plays a key role in gastrointestinal tract inflammation.
In conclusion, our studies demonstrated that colonic miR-141 was significantly reduced predominantly in the epithelial cells of the inflamed colons from colitic mice and CD patients, resulting in the upregulation of CXCL12β/total-CXCL12 in the colon. This pathway of miR-141 targeting CXCL12β has an important role in the development of intestinal inflammation, likely by influencing inflammatory cell trafficking into the inflamed sites. Inhibiting colonic CXCL12β expression and blocking immune cell recruitment with miRNA represents a promising approach that may be valuable for the treatment of CD.
Acknowledgments
We thank Dr Hongjie Zhang and Xin Dai for their assistance with patient tissue collection. We also thank Dr Zheng-Gang Liu from National Cancer Institute for his excellent advice.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
ZH and TS contributed equally.
-
Contributors ZH performed the experiment and wrote the manuscript; TS, QZ, SS, RZ and HS performed the experiment; LD performed the statistical analysis; CZ and KZ designed the experiment; JC and JZ designed the experiment and wrote the manuscript.
-
Funding This work was supported by the National Science Fund for Distinguished Young Scholars (81025019), the National Basic Research Program of China (2012CB517603), the National Natural Science Foundation of China (J1103512, J1210026, 31070722, 31271013, 31071232, 31170751, 31200695, 51173076, 91129712 and 81102489), the Key Project of the Chinese Ministry of Education (108059), the PhD Programs Foundation of the Ministry of Education of China (20100091120020) and the Scholarship Award for Excellent Doctoral Student granted by Ministry of Education.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval The local ethics committee.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement We are happy to share our data. All the reagents used are commercially available.
Linked Articles
- Commentary