Article Text

Abstract
Objective Surgical interventions that prevent nutrient exposure to the duodenum are among the most successful treatments for obesity and diabetes. However, these interventions are highly invasive, irreversible and often carry significant risk. The duodenal-endoluminal sleeve (DES) is a flexible tube that acts as a barrier to nutrient-tissue interaction along the duodenum. We implanted this device in Zucker Diabetic Fatty (ZDF) rats to gain greater understanding of duodenal nutrient exclusion on glucose homeostasis.
Design ZDF rats were randomised to four groups: Naive, sham ad libitum, sham pair-fed, and DES implanted. Food intake, body weight (BW) and body composition were measured for 28 days postoperatively. Glucose, lipid and bile acid metabolism were evaluated, as well as histological assessment of the upper intestine.
Results DES implantation induced a sustained decrease in BW throughout the study that was matched by pair-fed sham animals. Decreased BW resulted from loss of fat, but not lean mass. DES rats were also found to be more glucose tolerant than either ad libitum-fed or pair-fed sham controls, suggesting fat mass independent metabolic benefits. DES also reduced circulating triglyceride and glycerol levels while increasing circulating bile acids. Interestingly, DES stimulated a considerable increase in villus length throughout the upper intestine, which may contribute to metabolic improvements.
Conclusions Our preclinical results validate DES as a promising therapeutic approach to diabetes and obesity, which offers reversibility, low risk, low invasiveness and triple benefits including fat mass loss, glucose and lipid metabolism improvement which mechanistically may involve increased villus growth in the upper gut.
- BASIC SCIENCES
- ENDOCRINE HORMONES
- GASTROINTESTINAL ENDOCRINOLOGY
- OBESITY SURGERY
- OBESITY
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
Exclusion and/or altered presentation of nutrients in the duodenum is a key component of multiple bariatric interventions, including Roux-en-Y gastric bypass (RYGB).
-
Circulating bile acids are elevated after, and associated with the beneficial aspects of, RYGB.
-
Clinical and preclinical studies using duodenal-endoluminal sleeve (DES) that suggest reduced adiposity and improved glucose homeostasis.
What are the new findings?
-
DES implantation stimulates reduced body weight and fat mass in a rodent model of T2D.
-
DES-associated improvements in glucose and lipid homeostasis are fat-mass independent.
-
Profound changes in gut morphology and bile-acid dynamics may play an important role in the beneficial metabolic effects.
How might it impact on clinical practice in the foreseeable future?
-
These results validate DES as a promising therapeutic approach to diabetes and obesity, which offers reversibility, low risk and low invasiveness.
Introduction
Increasing obesity rates over the past three decades have resulted in a rapidly increasing incidence of metabolic disturbances, such as hypertension, dyslipidemia and type-2 diabetes (T2D). Conventional and available pharmaceutical therapies have been found to be only mildly efficacious,1 leaving surgical intervention as the most effective and durable therapy for severe obesity.2 Several bariatric interventions are currently used to produce sustained weight loss and treat obesity-related comorbidities. Of these, Roux-en-Y Gastric Bypass (RYGB) stimulates a considerable and sustained weight-loss in many obese patients that, perhaps more importantly, is accompanied by profound reversal of the diabetic state.3
Although at least some of the beneficial effects of RYGB on glucose control are secondary to reduced body weight (BW), dramatic changes in nutrient presentation (or lack thereof) and gut hormone secretion may also contribute to these effects.4 Clinical studies have identified changes in insulin-like growth factor 1, leptin, adrenocorticotropic hormone, ghrelin, peptide YY (PYY), and glucagon-like peptide-1 (GLP-1)5–7 after RYGB. These humoral changes are absent in weight loss attributed to either restrictive bariatric procedures or hypocaloric diet.8 ,9 While it is likely to be a component of the overall effect, humoral reprogramming alone is unlikely to account for the robust effects of RYGB on such a wide range of obesity-related comorbidities. Consistent with this hypothesis, is the observation that RYGB profoundly alters circulating bile-acid levels10 ,11 and gut microbiota,12 both of which may contribute to the beneficial effects. Taken together, these observations highlight the complex nature of RYGB, and provide multiple potential mechanisms for its effect on glucose metabolism.
One possible mechanism underlying the various effects of RYGB is the exclusion and/or altered presentation of nutrients in the duodenum. Accumulating evidence indicates that duodenal nutrient exclusion may be an important contributor to the metabolic benefits of RYGB.13–15 This observation is bolstered by studies in both rodents and man using duodenal, barrier-endolumenal sleeves (DES) that suggest reduced adiposity and improved glucose homeostasis.16–18 This procedure has been reported to represent a reversible therapy for the treatment of glucose intolerance, with the placement and retrieval of this device being accomplished via endoscopy. However, the mechanisms of action for DES, and the specific role of fat mass (FM) in this effect, remain to be elucidated.
The aforementioned data supports a robust association between duodenal nutrient exclusion and improved glucose metabolism. This hypothesis is supported by observations linking intestinal nutrient sensing to the central regulation of glucose and energy metabolism.19 ,20 In addition to their role as fuel, ingested nutrients also signal to the central nervous system (CNS) inhibiting further intake, and allowing for the regulation of energy balance.21 In this study, we used diet-independent rodent models of diabetes (Zucker Diabetic fa/fa (ZDF) rats) to gain mechanistic insight into the effect of duodenal nutrient exclusion on glucose tolerance. These studies identified a previously unknown FM-independent enhancement of glucose homeostasis in DES rats. Furthermore, we discovered that this enhancement is associated with increased plasma bile-acid levels and upper intestinal villus length. The studies described herein open up novel perspectives toward a more mechanistic understanding of the procedure's metabolic benefits.
Materials and methods
Animals
Ten-week-old, male ZDF rats from Charles Rivers (Wilmington, Massachusetts, USA) were individually housed and maintained on a 12/12 h light/dark cycle (lights off at 19:00 h) at 25°C and 50–60% humidity. Rats were provided ad libitum access to water and a standard, chow diet (Teklad LM-485, 5.6% fat). Before surgical intervention, all groups were matched for BW and FM. All procedures were approved by the University of Cincinnati Institutional Animal Care and Use Committee.
Duodenal-endoluminal sleeve implant
DES were created from silicone with a wall thickness of 0.1143 mm and outer diameter of 5 mm (FP Solutions Group, Exeter, New Hampshire, USA). DES contained a small cone region (10 mm diameter×10 mm long) and a 110 mm tail (figure 1A). DES implant was performed in isoflurane-anaesthetised rats as previously described.16 Briefly, chow was replaced with a liquid diet (Ensure Plus; 1.41 kcal/g; 29% fat; Abbott Nutrition, Columbus, Ohio, USA) 3 d prior to surgery. Rats were fasted for 16 h immediately prior to the procedure to empty stomach/intestinal contents. Laparotomy was followed by isolation of the stomach and the small intestine at the level of the ligament of Treitz. Two enterotomies were introduced, one 10–15 mm proximal to the pylorus and the second immediately distal to the ligament of Treitz. A rubber catheter was introduced via the duodenotomy and threaded through the duodenum to the stomach, in a retrograde manner. The DES was tied to the catheter and pulled into place by removal of the catheter. The cone of the DES was sutured into the antrum of the stomach via 4–6 ‘U-sutures’ using 6-0 Prolene, non-absorbable suture (Ethicon Endo-surgery, Somerville, New Jersey, USA) and anchored to a surgical pledget placed on the surface of the stomach. The enterotomies were then closed with a running 6-0 (antrotomy) or 8-0 (duodenotomy) Prolene non-absorbable suture (Ethicon). The stomach and upper intestine were then reintegrated into the peritoneal cavity and the abdominal wall and skin were closed in layers using a running suture and a running subcuticular suture, respectively. When implanted, this sleeve prevented nutrient-luminal exposure from the antrum of the stomach to the ligament of Treitz (figure 1B). The sham operation involved laparotomy, duodenotomy, antrotomy and passage of the catheter. All incisions were then repaired as in the DES procedure.
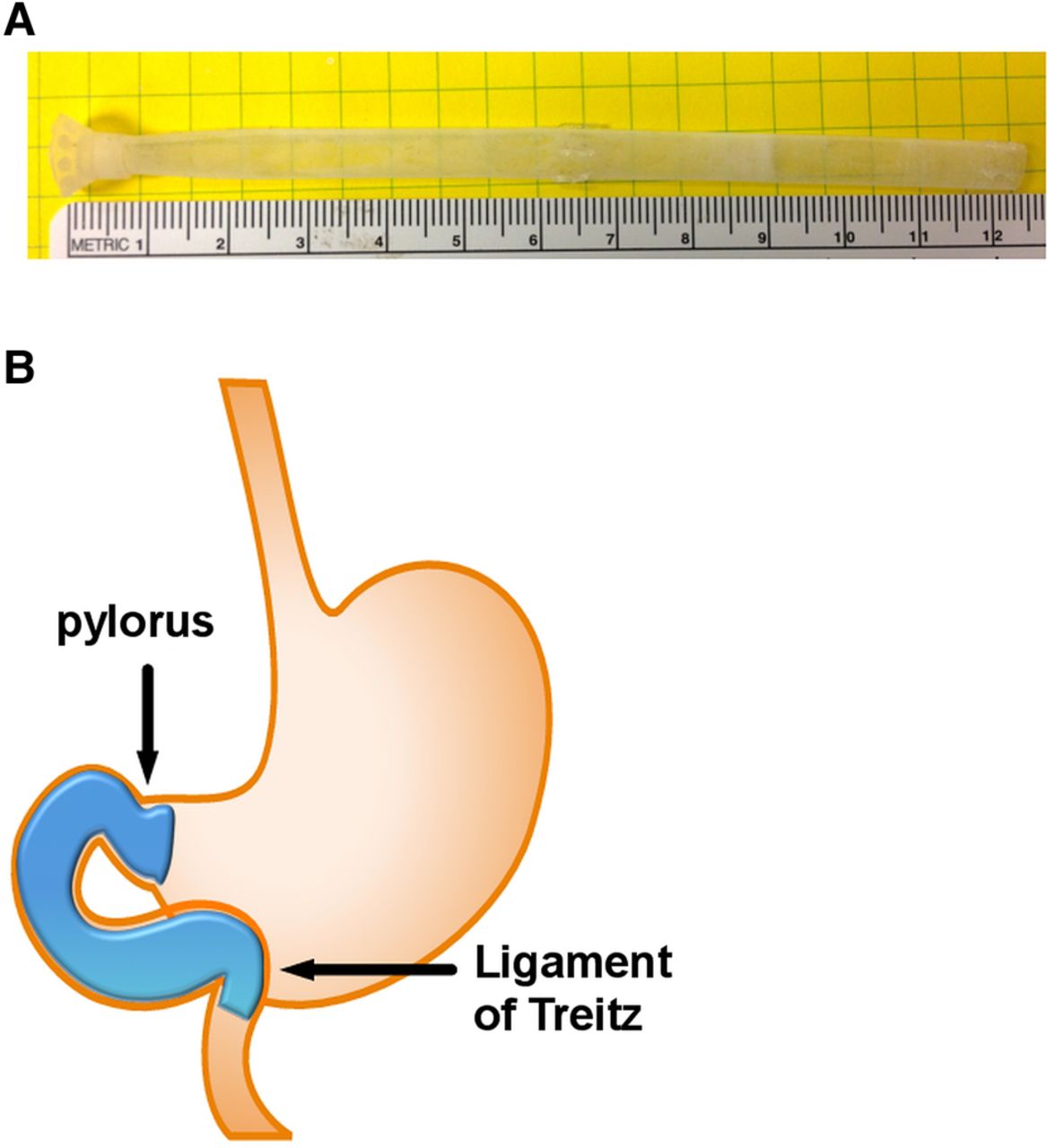
Characteristics of the duodenal-endoluminal sleeve: duodenal-endoluminal sleeve dimensions (A) and schematic of anatomical placement (B).
Postoperative care
In all surgical groups, liquid diet was continued for 120 h postoperatively and chow was reintroduced during the last 24 h. Subcutaneous injections of Metacam (0.25 mg/100 g BW once daily for 4 days), gentamicin (0.8 mg/100 g BW on the day of surgery), Buprenex (0.3 mL 2× per day for 5 days), and warm saline (10 and 5 mL 2× per day for days 0–3 and 4–5, respectively) were given to all postoperative rats. A wire grate was used for 72 h postoperatively to prevent rats from eating their bedding.
Body composition
Whole-body composition (fat and lean mass) was measured before the implant, as well as 14 and 27 days postoperatively using non-invasive nuclear magnetic resonance (NMR) technology (EchoMRI, Houston Texas, USA). The study was terminated following the final body composition measurement.
Glucose tolerance tests
Intraperitoneal (ip) and mixed-meal (mm) glucose-tolerance tests were performed by injection of glucose (20% w/v d-glucose, Sigma, in 0.9% w/v saline) or gavage of Ensure Plus (Abbott Nutrition, Columbus, Ohio, USA), respectively, after 6 h fasting. Blood glucose was sampled immediately before and 15, 30, 60 and 120 min after glucose administration. Plasma samples were collected immediately before and 15, 30 and 60 min after glucose challenge in protease inhibitor cocktail (Halt Protease and Phosphatase Inhibitor (Thermo Scientific, Logan UT) 4.65 g EDTA, and 40 000 U heparin in 50 mL saline). Plasma samples in the sham-pf groups were collected before, 15 and 60 min after glucose challenge. Electrochemiluminescence assay was used to measure insulin, glucagon, active and total GLP-1(7–36) (Meso Scale Discovery, Gaithersburg, Maryland, USA).
Intestinal histology
Duodenum, jejunum and ileum were harvested out of each rat at time of sacrifice. The 7 cm immediately proximal to the colon was removed from each rat. The most proximal 2 cm was placed in formalin and used for histological studies. The remaining 5 cm was split lengthwise and half was used to isolate RNA for the expression studies. Histological samples were immersed in 10% buffered formalin and submitted to the department of pathology at Cincinnati Children's Hospital Medical Center for H&E staining. Sections were examined using light microscopy and villus length (10 villi/section from 3 to 5 rats) was determined from images at identical magnification using ImageJ software (NIH).
Bile-acid quantification
Total serum bile acids were assayed using the 3α-hydroxysteroid dehydrogenase method (BQ Kits, San Diego, California, USA). The reaction was measured at 540 nm.
Gene expression
RNA was isolated using the RNeasy Lipid Mini-Kit (Qiagen, Valencia, California, USA) according to the manufacturer's instructions. cDNA was synthesised by reverse transcription PCR using SuperScriptIII, DNase treatment and anti-RNase treatment (Invitrogen, Carlsbad, California, USA). Single gene qPCR was performed according to the manufacturer's instructions on a 7900 real-time PCR machine (Applied Biosystems, Foster City, California, USA). Data were normalised to the housekeeping gene 18 s and expressed as fold change.
Statistical analysis
All data are represented as mean and SE of the mean (SEM). One-way and two-way analysis of variance (ANOVA) with Tukey's multiple comparison post-test were performed, where appropriate, using GraphPad Prism software (San Diego, California, USA) In all cases, statistical significance was assumed when p<0.05.
Results
DES alters body composition and energy balance
To test the metabolic effects of duodenal nutrient exclusion we implanted duodenal-endoluminal barrier sleeves (DES) similar to those described by Aguirre et al16 in ZDF rats. Unlike the previously described DES, our devices contained a cone that allowed them to be anchored in the distal portion of the antrum (figure 1A), thus generating a barrier from the gastric antrum to the ligament of Treitz (figure 1B). DES implantation decreased BW over the initial 5 d postoperative period compared to naive or sham rats (figure 2A). This weight loss was followed by a gradual increase in BW that matched the sham rats. Consistent with the initial decrease in BW, DES rats had reduced food intake over the initial 9 days of the study, with a subsequent increase in intake similar to that of naive or sham rats (figure 2B). Food intake in sham rats was also reduced over the initial postoperative period, but returned to naive-level intake on day 9 (figure 2B). In addition to the naive, sham and DES groups, an independent group of sham rats was established and pair-fed to the DES rats (figure 2B). Pair-fed, sham rats displayed similar BW (figure 2A) as compared to that observed in DES rats.

Duodenal-endoluminal sleeve (DES) effects on body weight (BW) and composition. (A) BW (% of preoperative BW), (B) daily food intake, (C) absolute fat mass (D) relative fat mass, (E) absolute lean mass and (F) relative lean mass in Zucker Diabetic fa/fa rats rats following DES implant or sham procedures. All data represented as mean±SEM of n=8–17 rats. *p<0.05, **p<0.01, and ***p<0.001; *vs sham and # vs naive.
To determine the effect of duodenal nutrient exclusion on body composition, rats from all groups were analysed for fat and lean mass on the day of the surgery and at 14 and 27 days after surgery. As with BW, all rats were well matched on day 0. DES rats displayed a suppression of absolute and relative FM accrual throughout the study (figure 2C,D). DES rats also displayed suppressed lean mass growth, while absolute lean mass increased in all other groups (figure 2E). However, in terms of the relative lean mass, DES rats displayed a leaner phenotype than any other group (figure 2F). Consistent with the BW response, pair-fed sham rats were similar to DES rats regarding lean mass and FM on day 14 (figure 2C–F). However, by day 27 there was a small but significant accrual of FM in these rats that was not observed in the DES rats (figure 2C,D).
DES improves glucose homeostasis
To identify the effect of duodenal nutrient exclusion on glucose homeostasis, we assessed several components of glucose regulation in our model. Naive rats displayed fasting hyperglycaemia on day 14, however, blood glucose was reduced in all surgical groups as compared to naive rats (figure 3A). While all surgical groups maintained euglycaemia, this regulation was achieved with considerably lower circulating insulin in DES and pair-fed rats (figure 3B). To further elucidate the role of duodenal nutrient exclusion in glucose homeostasis, glucose tolerance was assessed by ip glucose challenge (1 g/kg BW). This challenge resulted in pronounced glucose intolerance in naive rats as assessed by glucose excursion (figure 3C). Conversely, DES rats displayed enhanced glucose tolerance, with significantly decreased glucose excursions at 30, 60 and 120 min (figure 3C). Analysis of area under the curve of these excursions confirmed an enhancement in glucose tolerance in the DES group, with pair-fed rats displaying similar improvements (figure 3D). In this analysis, sham and naive rats were equally intolerant (figure 3D). Together, these data suggest that DES enhances glucose tolerance in ZDF rats, and these improvements are likely to be due to the reduction in diet-induced obesity.
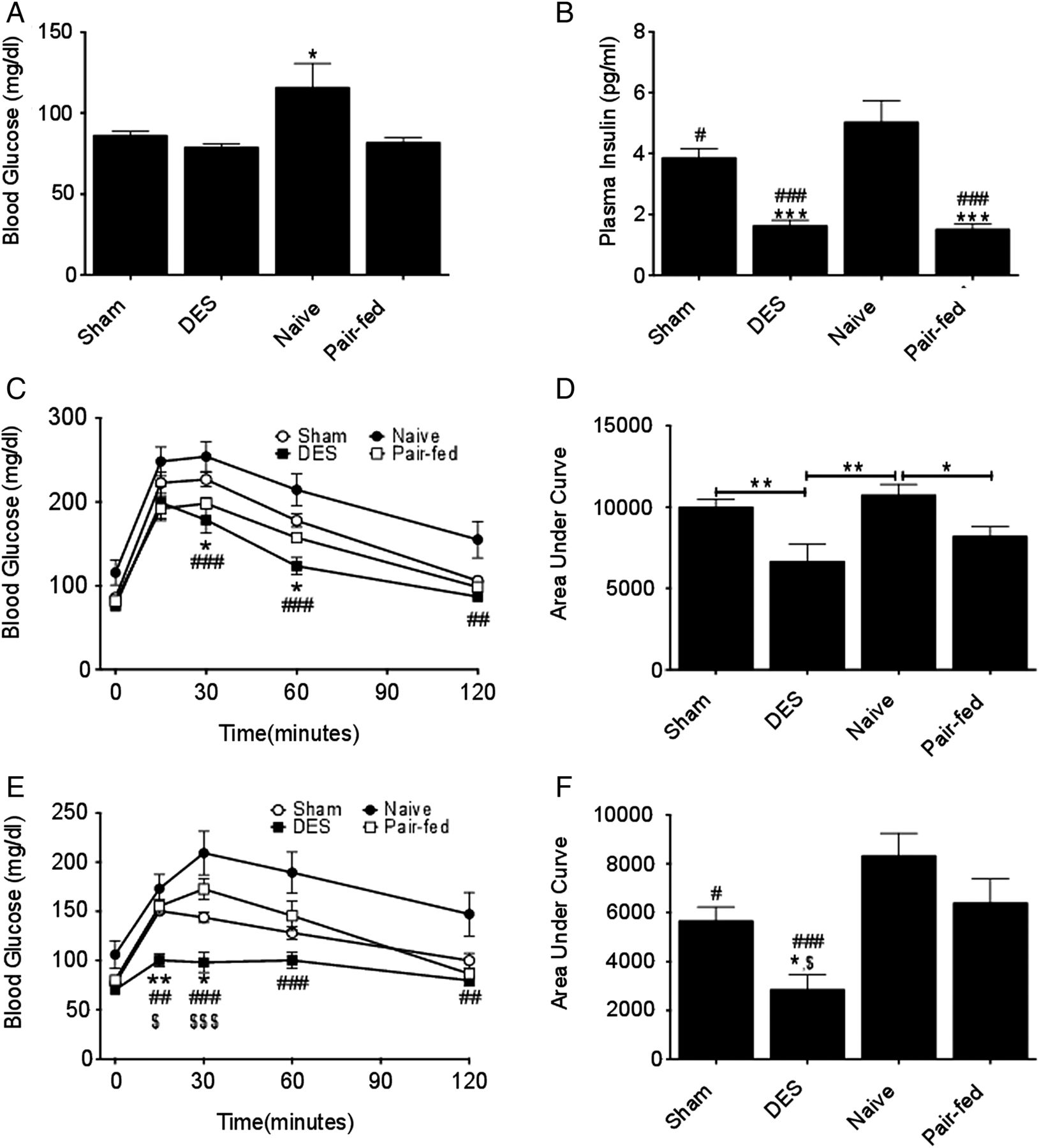
Duodenal-endoluminal sleeve (DES) improves glucose metabolism: fasting blood (A) glucose and (B) insulin on d 14. Glucose tolerance (C) as (assessed by 1.0 g/kg i.p. glucose challenge) and area under the curve analysis (D) following 4 h fast on d 14. Glucose tolerance (E) as (assessed by 2.5 g/kg oral glucose challenge in mixed meal) and area under the curve analysis (F) following 4 h fast on d 22. All data represented as mean±SEM of n=8–17 rats. *p<0.05, **p<0.01, and ***p<0.001; *vs sham, # vs naive, and $ vs pair-fed.
While our findings on the effect of DES on glucose tolerance are consistent with prior reports,16 the method of glucose challenge largely ignores the acute function of the barrier device and, instead, focuses on its long-term effects (FM reduction). To evaluate the effect of DES implantation on an ingested meal, we challenged the rats with an oral mixed meal. A liquid, mixed-meal diet (Ensure plus), was supplemented with 100 mg of d-glucose per mL diet. Thus, when each rat was delivered a flat dose of 5 mL on day 22, the average glucose dose was approximately 2.5 g/kg BW. This challenge elucidated similar excursion curves to those obtained with the ip challenge. However, under these conditions, the effect of the DES was heightened and significantly improved as compared with all other groups (figure 3E), highlighting the effects of duodenal nutrient exclusion on glucose tolerance. Likewise, the area under the curve of these excursions revealed enhanced glucose tolerance in DES rats as compared with all other groups (figure 3F). Taken together, these data suggest that duodenal nutrient exclusion profoundly enhances global and postprandial, glucose metabolism in a rat model of T2D (the ZDF rat).
DES stimulates GLP-1 secretion
The enhanced action of DES following oral glucose load compared to that observed with ip challenge suggests a possibly improved incretin effect triggered by duodenal nutrient exclusion. To test this hypothesis, we assessed humoral response to the mixed-meal challenge in these rats. Fasting insulin levels were suppressed in DES and pair-fed rats. Mixed-meal challenge in DES and pair-fed rats resulted in a plasma insulin peak at 15 min and overall lower insulin excursion as compared with the insulin peak in sham rats (figure 4A). While naive rats displayed intermediate plasma insulin at 15 min as compared to sham and DES rats, their excursion increased throughout the challenge (figure 4A), consistent with an impaired response to the glucose load. The 15 min insulin response (change from baseline plasma insulin) was similar in all groups, with the exception of naive rats (figure 4B), suggesting that the insulin secretion was appropriate to the glucose load. Although plasma insulin levels were suppressed in DES rats, active GLP-1 concentrations were elevated 60 min after a mixed meal (figure 4C). An increased area under the curve was also observed in DES rats as compared to all other groups (figure 4D). Consistent with decreased fasting plasma insulin, plasma glucagon was elevated prior to the mixed meal and similar to other groups thereafter (figure 4D). Area under the curve analysis of this test displayed a significant elevation in the DES rats as compared with the sham-pf controls, with a similar trend as compared with naive and sham rats. Together, these data show that DES rats exhibit enhanced glucose homeostasis associated with increased GLP-1 and glucagon, but decreased insulin.

Duodenal-endoluminal sleeve (DES) effects on hormone levels: plasma insulin excursion (A) and relative change (B) during mixed-meal challenge on day 22. Plasma GLP-1 (C) and glucagon (D) excursions during mixed-meal challenge on day 22. All data represented as mean±SEM of n=8–17 rats. *p<0.05, **p<0.01, and ***p<0.001; * vs sham, # vs naive, and $ vs pair-fed.
DES alters bile-acid homeostasis
The presence of a barrier device spanning the region from the gastric antrum to the ligament of Treitz will alter the site of bile-chyme interaction and possibly disrupt normal bile-acid dynamics. To test the hypothesis that DES would modify bile-acid homeostasis, we first assessed plasma bile-acid levels. Fasting bile acids were elevated in DES rats, while all other groups displayed similar plasma concentrations (figure 5A). Consistent with this elevation of circulating bile acids, liver expression of the bile-acid synthesis enzyme, sterol 27-hydroxylase (Cyp27a1), was suppressed, and cholesterol 7 α-hydroxylase (Cyp7a1) expression showed a similar trend towards reduced levels (figure 5B). Liver expression of organic anion-transporting polypeptide 1 (Oatp1) was elevated in sham-pf rats only. Liver expression of the Na+-taurocholate cotransporting polypeptide, organic anion-transporting polypeptide 4 (Oatp4), and the bile salt export pump (Besp) were unchanged by the presence of the endolumenal barrier (figure 5B). To assess a possible increase in farnesoid X receptor (FXR) activity following increased bile acid levels, we next investigated the expression of FXR-target genes in the ileum. Expression of both organic solute transporter α (OSTα), and organic solute transporter β (OSTβ) was suppressed in DES rats, with a similar trend observed in the FXR-target genes fibroblast growth factor 15 (Fgf15) and small heterodimer partner (Shp) (figure 5C). Together, these data suggest that elevated circulating bile acids in DES rats suppresses hepatic bile-acid synthetic enzymes, but is not associated with increased expression of FXR targets in the ileum.
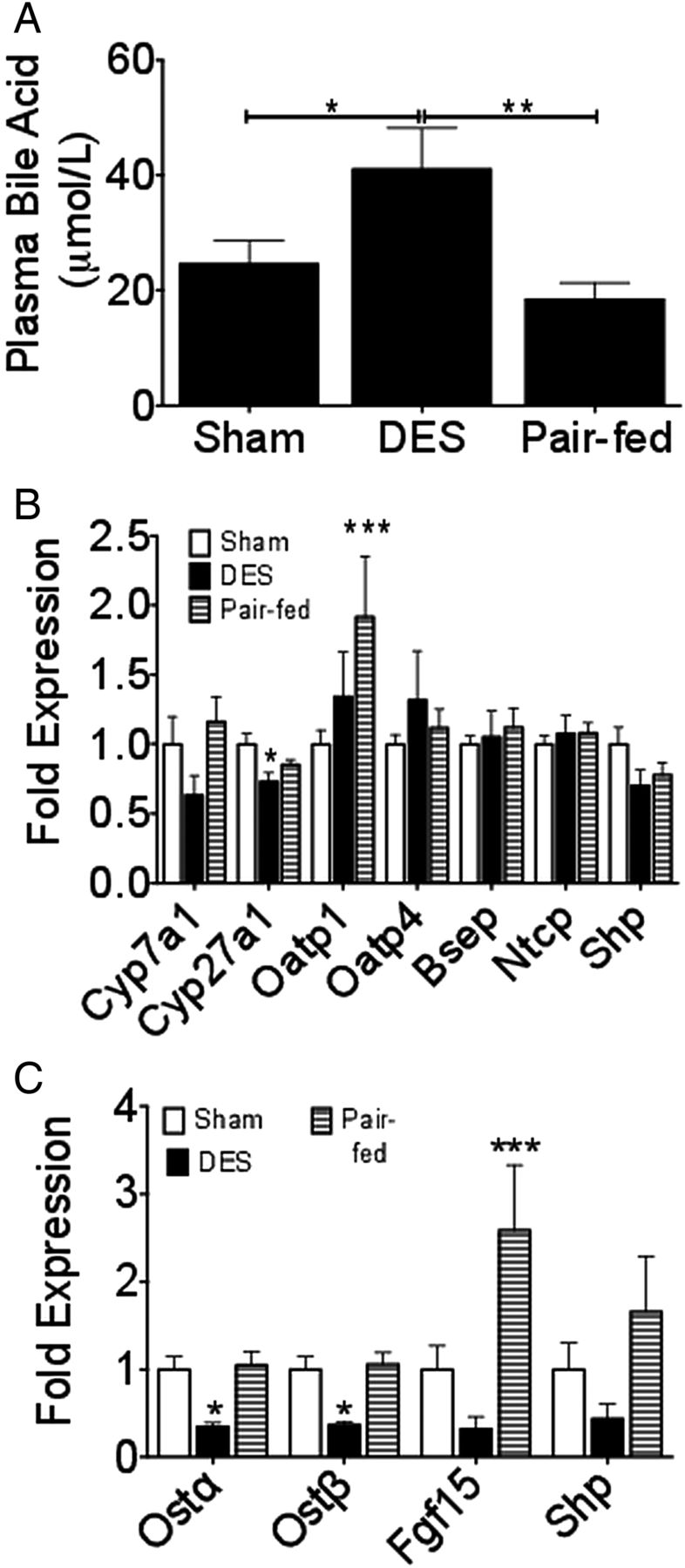
Duodenal-endoluminal sleeve (DES) alters bile-acid metabolism: (A) Plasma bile acids measured on d 27 following 16 h fast. Fold expression of liver Cyp7a1, Cyp27a1, and Na+-taurocholate cotransporting polypeptide (B) following 16 h fast. Fold expression of ileal OSTα and OSTβ (C) following 16 h fast. All data represented as mean±SEM of n=5–10 rats. *p<0.05, **p<0.01, and ***p<0.001; * vs sham.
DES stimulates upper intestinal villus growth
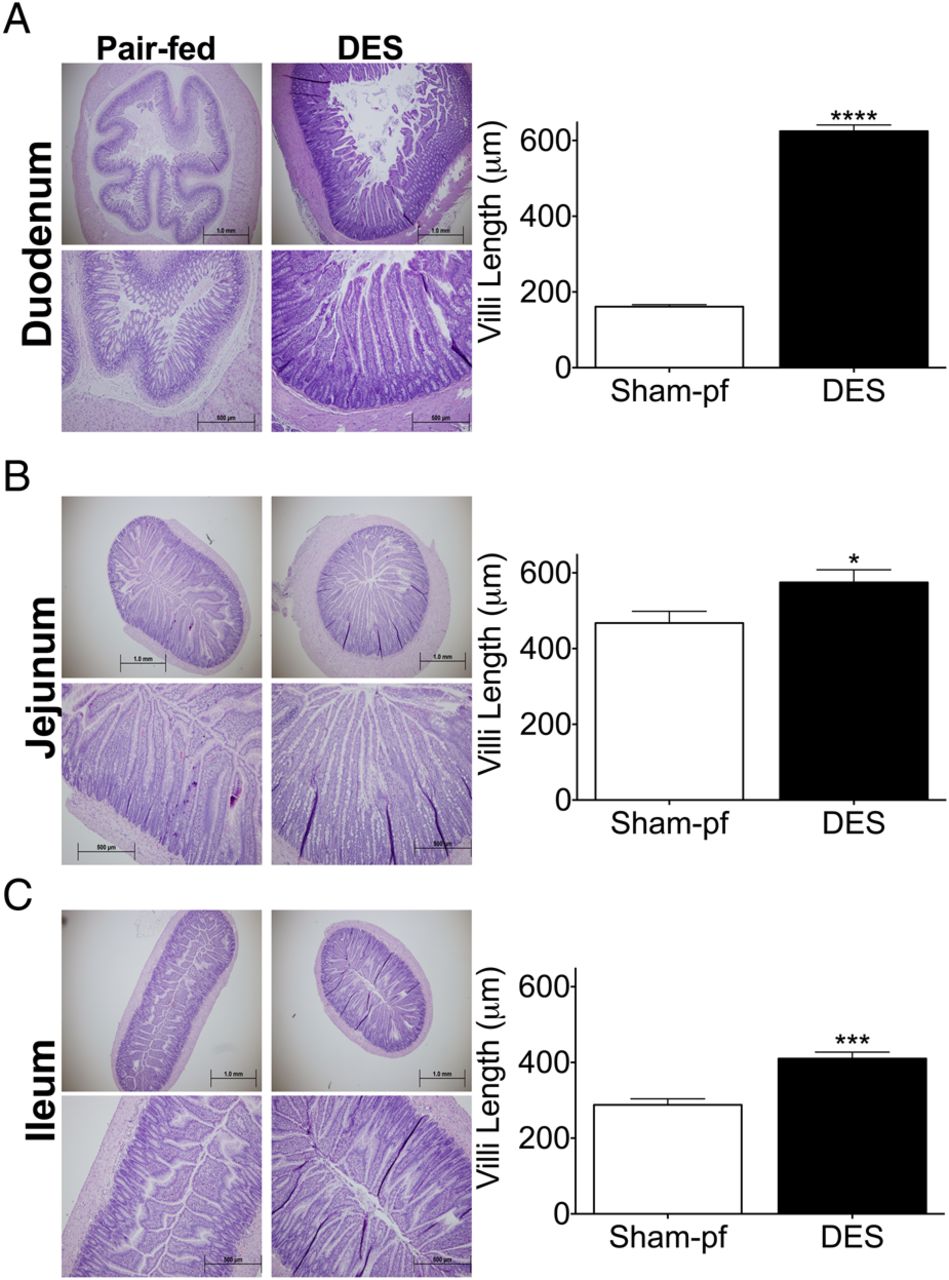
Bariatric interventions that alter bile-acid dynamics are often associated with profound structural changes of the upper intestine.22 ,23 To assess the effect of DES-induced duodenal nutrient exclusion on the upper intestine, matching sections of duodenum, jejunum and ileum were collected from DES and pair-fed rats at study termination. These groups allowed us to compare gut structure in rats matched for BW, FM and caloric intake over the course of the study. DES implantation stimulated profound morphological changes throughout the upper intestine including an overall increase in size. Qualitative observation of the duodenum uncovered a striking change in the basic morphology of the tissue. Specifically, we found a flattening of the submucosa resulting in a cylindrical and open configuration as compared to the highly invaginated lumen of the pair-fed rats (figure 6A). The flattening was accompanied by a profound increase in duodenal villus length, which was approximately 4.10-fold longer than pair-fed, sham rats (figure 6A). This effect was also observed in the jejunum (figure 6C,D) and ileum where villus length was increased by 1.34-fold and 1.60-fold, respectively. Taken together, these data suggest a profound morphological adaptation to the DES that begins in the duodenum and stimulates increased villus coverage, and presumably increased absorptive capacity in the jejunum and ileum.

Duodenal-endoluminal sleeve (DES) stimulates upper intestinal villus growth: Representative images (10× and 4×) and semiquantitative analysis of villus length of (A) duodenal, (B) jejunal, and (C) ileal sections from DES and pair-fed rats at termination of study. All data represented as mean±SEM of n=5–7 rats. ***p<0.001 vs pair-fed.
DES suppresses fasting and postprandial lipidemia
DES implant stimulates a leaner, more glucose-tolerant phenotype in this model of metabolic syndrome. To test the hypothesis that duodenal nutrient exclusion may also provide beneficial effects on metabolic syndrome-associated dyslipidemia, we assessed fasting and postprandial plasma lipid parameters in DES, ZDF rats. While fasting plasma cholesterol levels did not differ between groups, we observed an increase in the postprandial circulating cholesterol in pair-fed, sham rats that was prevented in the DES rats (figure 7A). Additional benefits were reflected by decreased circulating triglycerides in fasted DES rats compared with either sham (ad libitum) or pair-fed, sham controls (figure 7B). Furthermore, this effect was maintained in the postprandial state, where pair-fed, sham rats exhibited a further increase in circulating triglycerides (figure 7B). Analysis of sterol regulatory element-binding protein 1-c (SREBP) and fatty acid synthase gene expression in the liver reveal reductions of approximately 50% in the expression of both genes in DES rats compared with sham rats, regardless of feeding paradigm (figure 7C). Thus, duodenal nutrient exclusion appears to suppress hepatic lipogenic programming. To assess the effect of duodenal nutrient exclusion on global lipolysis, we measured circulating glycerol following a 16 h fast or 2 h after refeeding. We observed that fasting plasma glycerol levels were suppressed in DES rats as compared with either sham (ad libitum) or pair-fed, sham controls (figure 7D). In the postprandial state, all rats had decreased circulating plasma glycerol concentrations with the DES rats still maintaining a decreased level (figure 7D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Duodenal-endoluminal sleeve (DES) improves plasma-lipid metabolism: Circulating cholesterol (A), triglyceride (B), and glycerol (D) concentrations following 16 h fast or 2 h after refeeding on day 27. Fold expression of liver sterol regulatory element-binding protein 1-c and fatty acid synthase (C) following 16 h fast. All data represented as mean±SEM of n=5–10 rats. *p<0.05, **p<0.01, and ***p<0.001.
Discussion
The use of gut manipulations to treat obesity and glucose intolerance has emphasised the role of the upper intestine in glucose and lipid homeostasis, as well as in energy balance. While many of these bariatric interventions involve exclusion of nutrients from the upper intestine, a direct mechanistic link between duodenal nutrient exclusion and enhanced metabolic parameters following bariatric interventions, such as RYGB, are still the topics of intense investigation. In this study, we set out to test the hypothesis that duodenal nutrient exclusion, induced by a barrier device, would recapitulated some of the beneficial components of RYGB. Studies detailed herein suggest that DES implantation stimulates reduced BW and FM in a rodent model of T2D. Importantly, this manipulation was associated with an improvement in glucose and lipid homeostasis that was independent of the reduced FM. Furthermore, our results suggest that profound changes in gut morphology and bile-acid dynamics may play an important role in the beneficial metabolic changes.
DES alters body composition and energy balance
Exclusion and/or altered presentation of nutrients in the duodenum are a key component of RYGB, duodenal-jejunal bypass (DJB), and variations of these procedures. These procedures also lead to profound effects on BW and FM loss.24 Consistent with the effect observed in these surgical interventions, we observed a rapid decrease in BW associated with a transient decrease in food intake in rats with DES-induce duodenal nutrient exclusion. An important consideration of this intervention is that, unlike RYGB and DJB, our procedure preserves both gastric and intestinal tissue and orientation. Inclusion of a cohort of sham rats that were pair-fed to the DES group identified decreased food intake to be the primary driver of changes in body composition. The return to control-level food intake by day 12 was accompanied by a gradual increase in body and FM in DES rats, however, this growth was parallel to that of the ad libitum sham rats. Furthermore, the DES rats maintained decreased FM and a greater proportion of lean mass as compared to all other groups. Taken together, these data suggest that duodenal nutrient exclusion, independent of any other gut manipulation is sufficient to stimulate a more favourable energy balance in this rodent model of T2D.
DES improves glucose and lipid homeostasis
Similar to their effects on body composition and energy balance, procedures that exclude duodenal nutrient contact are often accompanied by rapid improvements in glucose homeostasis, which is often independent of weight loss.24 ,25 Furthermore, emerging data suggests that intestinal nutrient sensing is a central component in the regulation of glucose and energy metabolism.19 ,20 Our studies suggest that a similar argument can be made for DES. Specifically, our data show for the first time that DES rats are characterised by a robust enhancement in glucose tolerance. Intriguingly, this improvement is most robust, and even fat-mass independent, in the mixed meal challenge. Like RYGB, this enhanced glucose tolerance is associated with increased postprandial GLP-1 secretion.25 However, in the case of DES the increased tolerance and GLP-1 secretion was accompanied by decreased fasting and postprandial plasma insulin; while intriguingly, circulating glucagon was elevated. Thus, following DES implantation, ZDF rats are able to maintain fasting euglycaemia, and improved meal tolerance while secreting significantly less insulin. While this may seem counterintuitive, we feel that this result is indicative of changes in nutrient sensing in the proximal gut. In the ip glucose challenge, glucose is deposited directly to the islet stimulating a similar insulin secretion in the DES and pair-fed animals. However, macronutrients and micronutrients (glucose included) in the mixed-meal challenge are more highly regulated, beginning with absorption in the gut. This situation allows for systemic (possibly incretin and/or CNS) mediated regulation of glucose disposal. Thus, while the contribution of glucose-stimulated insulin secretion is similar in DES and pair-fed animals, the DES implant induces profound changes in the glucose homeostasis under these more physiological settings. Taken together, these data suggest that DES-induced duodenal nutrient exclusion stimulates improved glucose homeostasis that is independent of changes in body composition, and is characterised by enhanced insulin response.
As with the beneficial effects observed in glucose homeostasis, we found that duodenal nutrient exclusion reduced postprandial plasma lipid profile in weight-matched rats. The effect on circulating triglycerides was considerable and although the DES rats are still hyperlipidemic (approximately 340 mg/dL as compared to 39 or 235 mg/dL in a lean or obese Zucker rat,26 respectively), fasting plasma levels were reduced by almost half. In the context of reduced plasma insulin, this effect on circulating lipids supports enhanced insulin action in these rats. Intriguingly, postprandial cholesterol and triglyceride levels are elevated in the sham-pf rats. As such, an alternative interpretation of these data is that accelerated lipid adsorption as an adaptation to pair/meal feeding may be responsible for these effects.
DES alters bile acid homeostasis and stimulates upper intestinal villi growth
The robust increase in circulating bile acids after RYGB suggest a tantalising explanation for the phenomenon of enhanced glucose homeostasis after DES implant.27 In the context of our DES, a barrier device spanning the region from the gastric antrum to the ligament of Treitz will alter the site of bile-chyme interaction and possibly disrupt normal bile-acid dynamics. Consistent with this proposed disruption of bile-chyme mixing and similar to the effects observed in RYGB,27 we observed a 61% increase of circulating bile acids in ZDF rats after DES implant. This increase was independent of FM and caloric intake, as determined by comparison with the ad libitum and pair-fed sham groups. Consistent with the elevated plasma levels, expression of genes crucial to bile-acid synthesis was suppressed in the liver. Likewise, expression of bile-acid transporters in the ileum were suppressed following DES, suggesting a compensatory shift in response to the increase in circulating bile acids. While we observed a decrease in bile acid transporter expression, there may still be a greater number of receptors. This discrepancy between circulating bile acids and expression of bile acid transporters may be due to the increased surface area observed, or to a compensatory down-regulation of expression.
Changes in bile-acid dynamics27 and glucose homeostasis27 ,28 are associated with increased villus height in the common limb following RYGB28 and ileal transposition.22 ,28 The presence of changes in both glucose tolerance and plasma bile-acid concentrations led us to postulate that gut morphology may also be altered in DES. To normalise for any changes induced by the decreased BW or transient hypophagia, we assessed upper intestines of DES rats with respect to pair-fed, sham rats. Consistent with our hypothesis, we found increased villus length and surface area throughout the upper intestine. Intriguingly, these changes have been suggested to represent a shift from an absorptive to a secretory cell linage and an increase in ileal glucose uptake.28 These associated effects open the door to a reversible therapeutic approach involving stimulation of villus growth, by DES or even by bile-acid mimetics.
The current data characterise a reversible therapeutic approach that improves a number of metabolic parameters. These studies highlight the beneficial effects of duodenal nutrient exclusion in a rodent model of T2D. Specifically, we find that duodenal nutrient exclusion reduced BW and FM, enhanced glucose tolerance, and corrected dyslipidemia. Importantly, these data also offer insight into a possible bile-acid-mediated mechanism, uniting these divergent outcomes, and providing targets for future therapies.
Acknowledgments
We would like to thank Mouhamadoul Toure, Alfor Lewis and Kathi Smith of the University of Cincinnati for their surgical expertise in conducting the DES and sham surgeries.
References
Footnotes
-
Contributors KMHa and MHT were responsible for study conception and design, data analyses and interpretation, and drafting the article; OAL, KMHe, AM, JH, JB, CXY, YG, ML, NO, SA and CR, acquired data; TM, PTP, RK, DP-T and RJS advised study concept and critical revision of the article. MHT is the guarantor of this work, had full access to all the data, and takes full responsibility for the integrity of data and the accuracy of data analysis.
-
Funding This work was supported by NIH grants: R01 DK077975, Neuroendocrine Regulation of Adipocyte Metabolism (DPT), K08 DK084310-03 (RK), the University of Cincinnati Training Program in Neuroendocrinology of Homeostasis grant T32 DK059803 (KMH), and Ethicon endosurgery.
-
Competing interests MHT and DP-T have a collaborative association with Roche Research Laboratories pertaining to peptide-based therapeutics in metabolism. MHT receives research funds from and is a Consultant for Roche Pharmaceuticals. RJS is a Consultant for Ethicon Endo-Surgery, Novo Nordisk, Novartis, Angiochem, Zafgen, Takeda, Zealand & Eli Lilly; receives research support from Ethicon Endo-Surgery, Novo Nordisk, Ablaris, Zealand & Pfizer; is a paid speaker for Ethicon Endo-Surgery, Novo Nordisk, Merck, & Pfizer; and holds equity in Zafgen.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement Data is available for scientific sharing.