Article Text

Abstract
Aim—Cholangiocarcinoma can be cured by surgery, but only in a minority of cases. The activation of apoptosis is a major mode of action of chemotherapy and radiotherapy, which have limited benefit in the treatment of cholangiocarcinoma. The antiapoptotic members of the Bcl-2 protein family (Bcl-2, Bcl-XL, and Mcl-1) are important inhibitors of apoptosis, but have not been investigated extensively in cholangiocarcinoma.
Methods—The expression of Bcl-2, Bcl-XL, and Mcl-1 was investigated in normal biliary epithelium (17), biliary dysplasia (three), and invasive cholangiocarcinoma (51), in addition to three human cholangiocarcinoma cell lines, by immunohistochemistry and immunofluorescence.
Results—The expression of Bcl-2 was not detected in normal or malignant biliary tissue. In contrast, granular cytoplasmic Bcl-XL and Mcl-1 staining was found in 60–100% of cells in all normal, dysplastic, and malignant specimens, including the human cell lines examined in this study.
Conclusion—These findings indicate that Mcl-1 and Bcl-XL, but not Bcl-2, are involved in the survival of normal and neoplastic cells in the biliary tree. By prolonging survival through blocking apoptosis, these proteins might be reducing the efficacy of cytotoxic anticancer treatments in cholangiocarcinoma.
- cholangiocarcinoma
- apoptosis
- Bcl-2
- Mcl-2
- Bcl-XL
Statistics from Altmetric.com
Cholangiocarcinoma is the second most common primary malignancy of the liver. The aetiology remains unknown. Oxidative damage, oncogene activation, and impaired apoptosis have been suggested to play a part in the pathogenesis.1 Despite advances in diagnosis, surgery offers the only possible chance for long term survival.2–4 Resection rates are at best between 25–50%.5 The five year survival rate after curative resection is between 0% and 30%.6–8 The results of palliative chemotherapy and/or radiotherapy remain poor.9–12 Chemotherapy and radiotherapy used in the treatment of a wide variety of malignancies induces apoptosis in responsive cells. Failure to initiate apoptosis is a major factor limiting the efficacy of these treatments.13,14 The Bcl-2 protein family plays a central part in the control of apoptosis.15,16 Bcl-2, a 52 kDa protein, is the prototype of this family, and inhibits the induction of apoptosis. Bcl-XL, Mcl-1, Bcl-w, and A1 are other antiapoptotic members, whereas Bax, Bad, Bid, and Bcl-XS are pro-apoptotic. High concentrations of Bcl-2 or Bcl-XL affect the susceptibility of a cell to the induction of apoptosis by altering the ratio of death promoters to suppressors, providing tumour cells with a survival advantage, and permitting expansion of transformed cells harbouring mutations within their genome. The Bcl-2 protein and other family members target intracellular organelles such as the endoplasmic reticulum and the outer mitochondrial and nuclear membranes, where they modulate responses to diverse death stimuli. This subcellular location is important for function.17 Some members of the Bcl-2 protein family when bound to the mitochondria regulate the function of the large polymeric channel, permeability transition pore complex,18 located at the point of contact between the inner and the outer mitochondrial membranes.19 The opening of this pore, known as mitochondrial permeability transition, is associated with the release of apoptogenic proteins.20,21
Some cases of cholangiocarcinoma have been found to express Bcl-2.22–24 Studies involving a variety of haematological and several solid malignancies have identified a correlation between high Bcl-2 or Bcl-XL expression, poor patient response to treatment, and overall prognosis.25–27 Such observations parallel the findings in experimental models.28–32 No such correlation has been suggested or studied in cholangiocarcinoma.
Cholangiocarcinoma is multidrug and radioresistant. Failure to induce apoptosis after cytotoxic treatment could explain the poor response to treatment. Therefore, the expression of the Bcl-2, Bcl-XL, and Mcl-1 proteins was investigated in normal and malignant biliary epithelium.
Materials and methods
PATIENT SPECIMENS
Cholangiocarcinoma specimens (formalin fixed, paraffin wax embedded) from 51 patients who had undergone surgery over the past 20 years in our department were examined. All 51 cases had the diagnosis verified by a consultant pathologist before being included in our study, at the same time the blocks most representative for study were selected. Normal biliary epithelium tissue comprised a combination of resected normal gallbladders (six), common bile duct (one), and bile duct brushings collected during endoscopic retrograde cholangiopancreatography (ERCP) for benign disease (10). In addition, three cases of biliary dysplasia were examined. Blocks were either cut 5 μm thick for conventional morphology (haematoxylin and eosin stain) or 3 μm (sequential sections) for immunohistochemistry.
Brush and bile samples collected at ERCP were centrifuged at 900 ×g for five minutes, the cell pellet was then resuspended in RPMI medium (Sigma, Poole, Dorset, UK) and two drops of albumin added. Aliquots (100 μl) of the cell suspension were then mounted on a slide by cytocentrifugation at 91.5 ×g for two minutes. Slides were either routinely alcohol fixed and stained by the Papanicolau method or allowed to air dry before immersion in acetone at room temperature for immunohistochemistry.
Schwann cells, intraepithelial lymphocytes (small T cells), and peripancreatic lymph nodes present within resected specimens were used as internal positive controls, and tonsil slides (frozen tissue) provided additional positive controls.
HUMAN CELL LINES
Egi-1, Tfk-1, and Sk-ChA-1 are three well characterised anchorage dependent human cholangiocarcinoma cell lines derived from patients before exposure to chemotherapy or radiotherapy and before surgery. Egi-1 and Tfk-1 were commercially obtained (DSMZ, Braunschweig. Germany), whereas Sk-ChA-1 was a gift from Professor Knuth of the Ludwig Institute For Cancer Research (Postfach, 8024 Zurich, Switzerland).33,34
IMMUNOHISTOCHEMISTRY
Sequential tissue sections (3 μm) were mounted on 3-aminopropyltriethixysaline (APES; Sigma) coated slides. Archival samples were dewaxed by graduated washes in xylene, then dehydration in absolute alcohol. Endogenous peroxidase was blocked by bathing for 10 minutes in 3% H2O2, after which slides were washed in running water. Antigen retrieval was performed by microwaving (800 W, full power) in citrate buffer (pH 6–7) for 20 minutes. Slides were then bathed in water for two minutes. Protein blocking was achieved by incubation in 10% normal goat serum for 10 minutes.
Using a three step streptavidin–biotin method for Bcl-2 staining, slides were incubated with antihuman Bcl-2 primary monoclonal antibody (Dako, Ely, Cambridgeshire, UK) at a 1/50 dilution, or negative mouse IgG1 (Dako), at a 1/400 dilution, at room temperature for one hour. Secondary (1/100 dilution) and tertiary (1/100 dilution) layers were incubated for 30 minutes each (Dako ABC kit). For Bcl-XL and Mcl-1 staining the catalysed signal amplification system (CSA, K1500; Dako) was used. This method also incorporates streptavidin–biotin complex, but has additional steps made up of biotinylated tyramide signal enhancement (recommended by Dako).35 The system was adapted for rabbit primary antibody using the biotinylated antirabbit link (Dako). Antihuman Mcl-1 (Dako) and Bcl-XL (Autogen Bioclear, Calne, Wiltshire, UK) primary polyclonal antibodies were incubated with slides through a series of 15 minute steps at room temperature at a dilution of 1/1000 and 1/750, respectively. Normal rabbit immunoglobulin as the first antibody layer (serum) formed the negative control. To reduce non-specific background staining, 0.1% Tween 20 (wt/vol) and 0.1% bovine serum albumin (wt/vol) were added to Tris buffered saline (pH 7.6). This solution was used routinely to dilute the antibodies and wash the sections. In addition, the sections were bathed in the background reducing solution for five minutes between each step during the staining process. Slides were then developed using 3`3-diaminobenzidine tetrahydrochloride (DAB) for 10 minutes, followed by counterstaining with haematoxylin. Slides were examined by light microscopy and the degree and number of cells staining were assessed alongside the corresponding haematoxylin and eosin specimen by two independent observers as negative, mild (1+), moderate (2+), or intense (3+) positive staining.
FLUORESCENCE MICROSCOPY AND IMMUNOFLUORESCENCE ANALYSIS OF BCL-2, MCL-1, AND BCL-Xl
The adherent cholangiocarcinoma cell lines Tfk-1, Egi-1, and Sk-ChA-1 were trypsinised preconfluence using trypsin/EDTA (×1; Sigma). After washing in buffered saline, cells were fixed and permeabilised using the Dako intrastain kit. Fluorescein isothiocyanate (FITC) conjugated rabbit antihuman Mcl-1 and Bcl-xL polyclonal antibodies and FITC monoclonal antihuman Bcl-2 antibody (Dako) were incubated at a 1/100 dilution in the dark at room temperature for 15 minutes. Negative isotype controls (rabbit Ig) were used at a 1/100 dilution. Cells were mounted on slides and images captured using a Zeiss Axioskop fluorescent microscope in conjunction with Iplab spectrum image analysis software (version 3.1.1).
Results and Discussion
Cholangiocarcinoma cells and normal biliary epithelial cells examined did not express the Bcl-2 protein, despite positive Bcl-2 staining in control tissue and small intrahepatic regenerating bile ductules (table 1; fig 1). However, Bcl-XL and Mcl-1 were expressed in all cholangiocarcinoma specimens examined, including the three human derived cholangiocarcinoma cell lines (figs 2, 3C,D). In addition, benign biliary epithelial cells from both preserved tissue and fresh samples also expressed Bcl-XL and Mcl-1 (fig 3A,B). The distribution of the cytoplasmic staining at higher magnification was localised to intracellular organelles, such as the mitochondria; immunofluorescence demonstrated the distribution more clearly (fig 3C,D). Although a double staining immunohistochemical technique would be required to prove formally that both Bcl-XL and Mcl-1 are coexpressed in the same cell, the staining from sequential sections and cell lines for both antibodies strongly suggest that this is the case.
Expression of Bcl-2, Bcl-XL, and Mcl-1 in normal, dysplastic, and malignant biliary epithelium

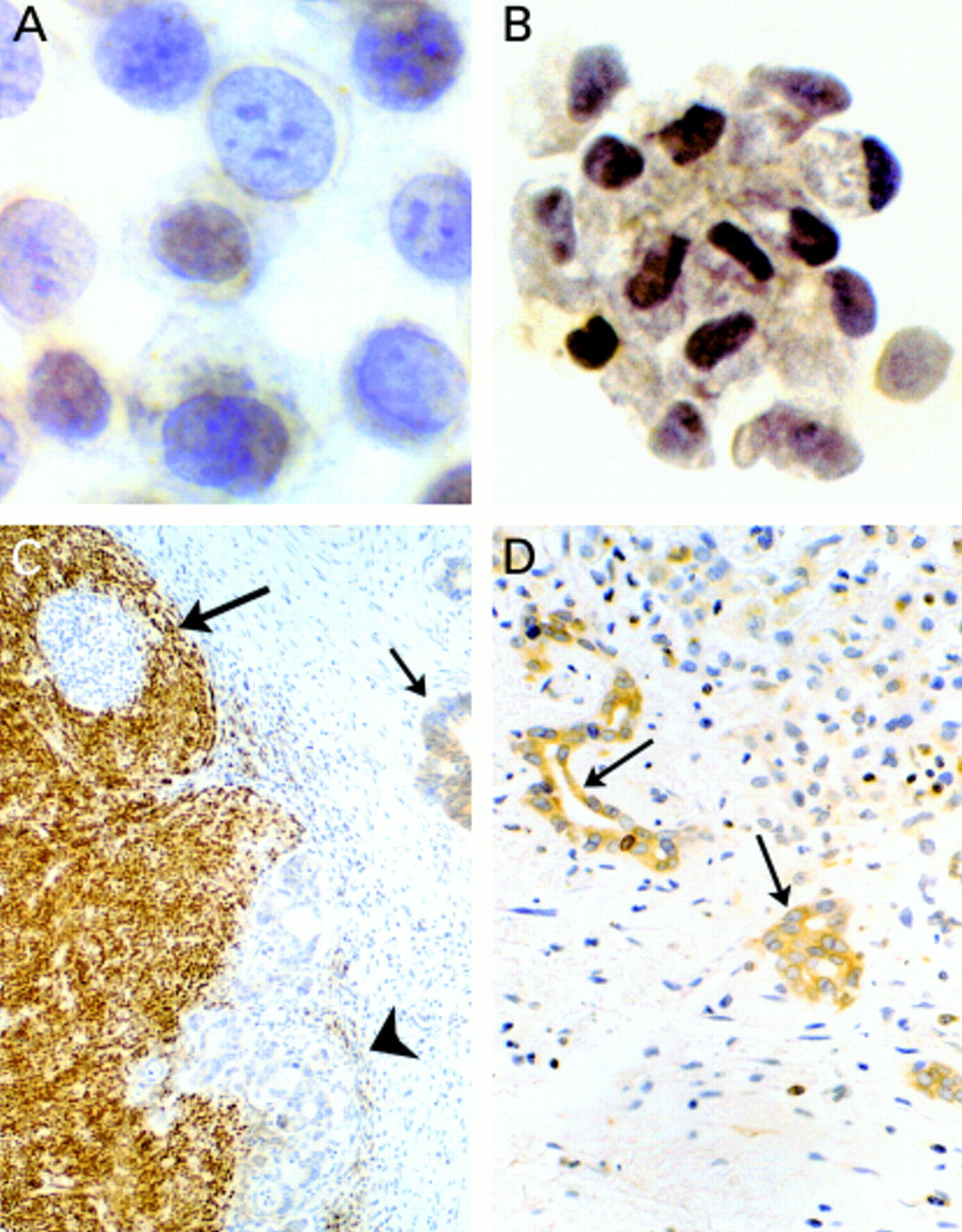
Bcl-2 protein expression in biliary epithelial cells. (A) The human cholangiocarcinoma cell line Egi-1 does not express Bcl-2. (B) Normal bile duct brush cytology cells stain negative for Bcl-2. (C) Cholangiocarcinoma cells located in the desmoplastic reaction (arrow) and within a lymph node (arrowhead) do not express Bcl-2. Positive Bcl-2 staining is clearly demonstrated in the internal positive control (lymph node). (D) Positive Bcl-2 staining is shown in cells of regenerating bile ductules.

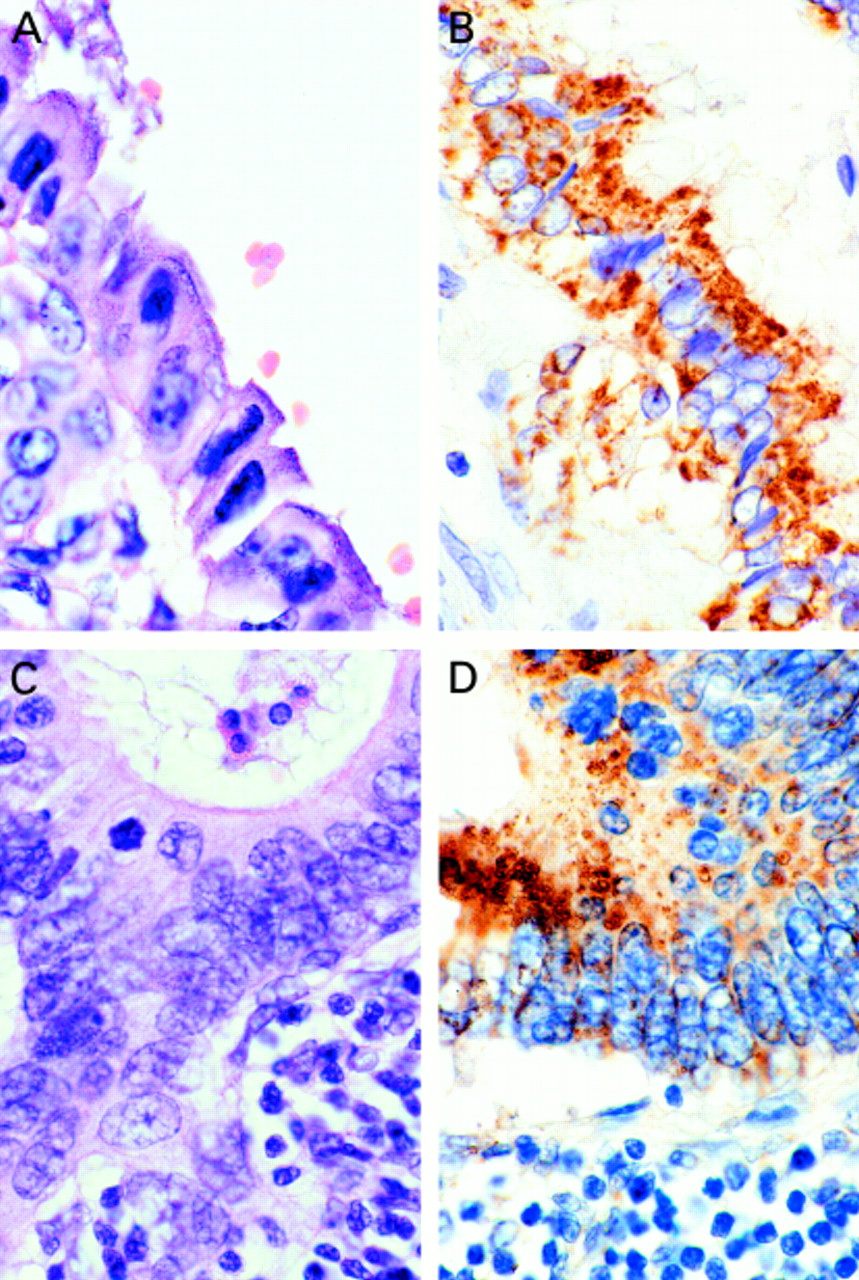
(A,C) Haematoxylin and eosin stain of cholangiocarcinoma. (B) Bcl-X L expression in cholangiocarcinoma. (D) Cholangiocarcinoma cells within a lymph node express Mcl-1; note the granular cytoplasmic pattern of staining for the Bcl-X L and Mcl-1 proteins.

{kind=link}
{kind=link}
{kind=link}
(A) Normal brush cytology cells and (B) the human cell line Tfk-1 also express Bcl-X L . Immunofluorescence images show (C) the Bcl-X L and (D) Mcl-1 proteins in Egi-1 localise to intracellular organelles, such as the mitochondria.
Most studies looking at the expression of Bcl-2 in cholangiocarcinoma report varying amounts of Bcl-2 protein expression in this disease.24 In contrast, a study looking at Bcl-2 protein expression in cholangiocarcinoma recently reported that none of the cholangiocarcinoma samples examined expressed the Bcl-2 protein.36 With Bcl-2 expression detected in the internal positive controls and in regenerating intrahepatic ductular epithelium, an observation reported previously,37 our inability to detect the of Bcl-2 protein is unlikely to be a result of poor antigen retrieval or low Bcl-2 antibody activity. There have been suggestions that in some cell types a reciprocal expression of survival proteins Bcl-2 and Bcl-XL occurs. Considering there are still many unanswered questions about the cellular control and preference of expression of the different Bcl-2 genes, this might explain the differences in reported findings. It may be that as we learn more about these proteins, reasons for the differences in expression may become clearer. The number of cholangiocarcinoma samples examined (archival tissue from 51 cases and three cell lines) makes this the largest study of Bcl-2 expression in cholangiocarcinoma. In combination with the findings of the recent study by Arora and colleagues,36 it seems that most cases of cholangiocarcinoma do not express immunohistochemically detectable amounts of Bcl-2 protein, and its function is probably carried out by other proteins, such as Bcl-XL. What is of more interest, however, is that the expression of antiapoptotic proteins appears to be a common feature in cholangiocarcinoma.
The expression of antiapoptotic (cell survival) proteins by normal biliary epithelial cells may be an adaptive response aimed at protecting against excessive cellular loss as a result of apoptosis caused by biliary toxins such as bile salts, xenobiotics, drugs, and heavy metals. Because the epithelial lining of the bile ducts is a monolayer, the balance of cell loss against cell production will require tight control to avoid excessive losses as a result of apoptosis, which could lead to ulceration, scarring, and fibrosis of bile ducts. The regulation of apoptosis within the biliary tree to maintain tissue homeostasis may be one function of biliary Bcl-XL and Mcl-1 proteins. A recent study found that skin keratinocytes that are constantly exposed to the cytotoxic effects of UV light were resistant in vitro to irradiation induced apoptosis through the upregulation of Bcl-XL protein expression.38 That the Bcl-XL and Mcl-1 proteins are playing a similar role in the biliary tree has yet to been shown. There are some researchers who have hypothesised that high concentrations of antiapoptotic proteins, such as the Bcl-2 protein, within the biliary tree play a key role in the development of cholangiocarcinoma by allowing cells that harbour growth promoting mutations, such as mutations of K-ras, c-erb-B-2, or c-myc, to survive by evading apoptosis, which would otherwise be triggered by an unstable genome.1
The role that antiapoptotic proteins play in the pathogenesis and the response of cholangiocarcinoma to cytotoxic treatment are unclear. It may be that studies looking into the effects on apoptosis sensitivities after antagonising Bcl-XL and/or Mcl-1 target function, directly or indirectly, may provide some evidence for their possible role in the resistance to anticancer treatment.
In conclusion, our study of Bcl-XL, Mcl-1, and Bcl-2 protein expression in benign and neoplastic biliary epithelium demonstrates that both normal and neoplastic cells coexpress the cell survival proteins Bcl-XL and Mcl-1, but not Bcl-2. Through inhibiting apoptosis, the Bcl-XL and Mcl-1 proteins expressed by cholangiocarcinoma may be contributing to the low efficacy of chemotherapy and radiotherapy in this disease.
Acknowledgments
This study was supported financially by the Liver Cancer Research Fund (RFUCSM grant BSAE to BD). We are grateful for the technical assistance of Miss L Garwood, Department of Cytopathology, Royal Free Hospital.