Article Text

Statistics from Altmetric.com
The viscera receive dual sensory innervation. The majority of visceral sensory fibres terminate in the spinal cord but sensory fibres contained in the vagus and pelvic nerves, which terminate in the brain stem and lumbosacral spinal cord, respectively, also innervate the same visceral organs. Figure 1 illustrates this unique pattern of innervation for the gastrointestinal tract, classically referred to as sympathetic and parasympathetic, but more appropriately designated by nerve name (for example, hypogastric nerve, pelvic nerve).
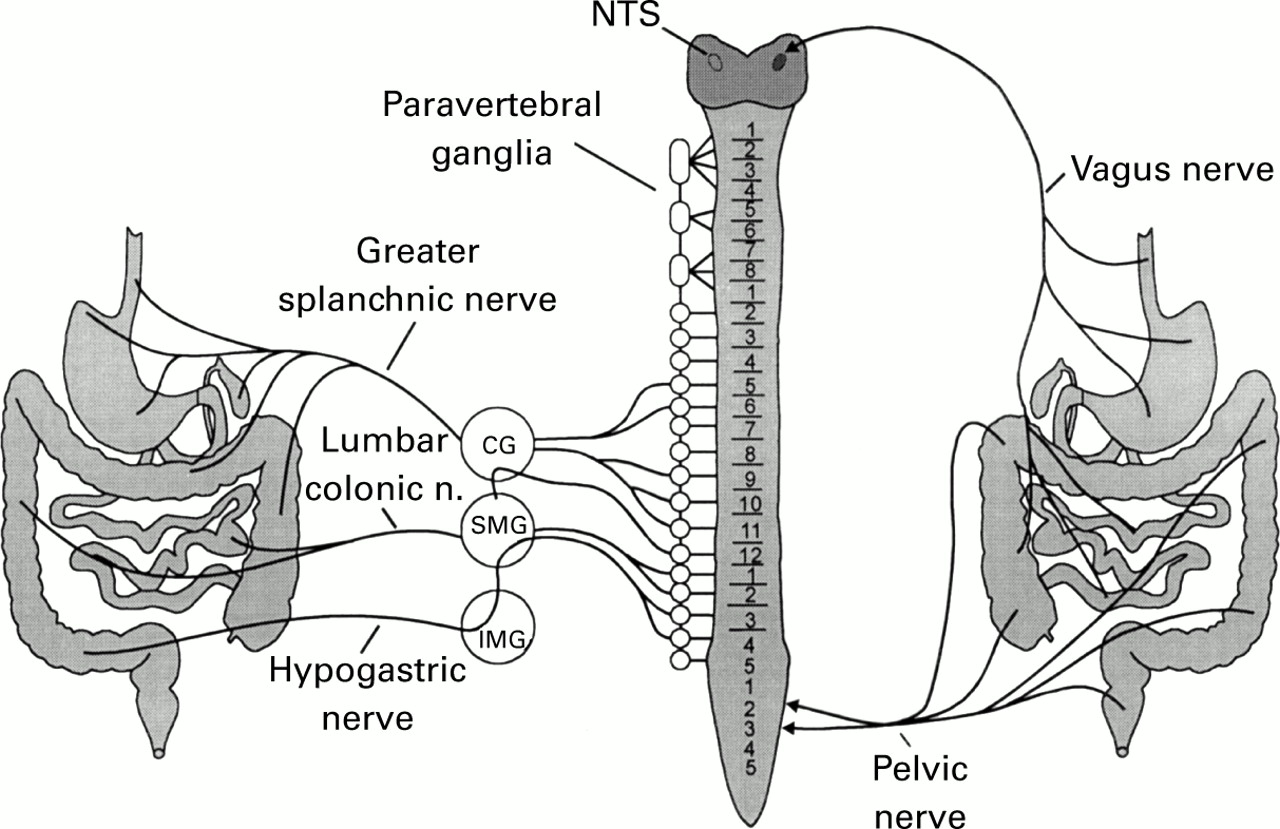
Representation of visceral sensory innervation of the gastrointestinal tract. The sensory innervation that anatomically exists in association with the sympathetic nervous system is shown on the left. These spinal visceral sensory fibres traverse both prevertebral (CG, coeliac ganglion, IMG, inferior mesenteric ganglion, SMG, superior mesenteric ganglion) and paravertebral ganglia en route to the spinal cord. Pelvic and vagus nerve innervation to the sacral spinal cord and brain stem, respectively, is shown on the right. (Modified from Sengupta and Gebhart.1)
Visceral sensory axons are almost exclusively thinly myelinated Aδ fibres and unmyelinated C fibres. Activity in most visceral sensory neurones, whether in vagal afferent fibres or spinal afferent fibres, does not reach consciousness. For example, there is regular input into the central nervous system from gastric and hepatic chemoreceptors, aortic baroreceptors, etc., that is not perceived.
Sensory innervation of the gastrointestinal tract involves all layers of a viscus (mucosa, muscle, and serosa) and visceral receptors exhibit chemosensitivity, thermosensitivity, and mechanosensitivity. Visceral receptors, in fact, are generally polymodal in character (that is, respond to multiple modalities of stimulation). The cell bodies of visceral afferent neurones, in common with the cell bodies of somatic afferent neurones, are located in dorsal root ganglia (except for the nodose ganglia which contain the cell bodies of vagal sensory neurones). However, the route visceral afferent neurones take to the spinal cord typically involves passage through or near prevertebral ganglia (where they can give off collateral axons to influence autonomic ganglion cell bodies and, accordingly, secretory and motor functions) and paravertebral ganglia (see fig 1).
In contrast with afferent fibres arising from somatic structures, the number of spinal visceral afferent fibres is estimated to be less than 10% of the total spinal afferent input from all sources. Some compensation for this relative paucity of visceral input is provided by the significantly greater rostrocaudal intraspinal spread of visceral afferent fibre terminals. Visceral C fibres have been found to have many more terminal swellings (suggestive of synapses) and terminal areas, and to exhibit rostrocaudal distribution over several spinal cord segments.
Experimentally, balloon distension is the most reliable stimulus for hollow visceral organs and is the most widely used stimulus in both human and non-human animal experiments.2 Accordingly, we know most about the response properties of mechanosensitive visceral afferent fibres, usually those innervating the muscle layer. Hollow viscera are innervated by two populations of mechanosensitive afferent fibres: a larger group (70–80%) of fibres have low thresholds for response and a smaller group (20–30%) of afferent fibres have high thresholds for response (for example, >30 mm Hg distending pressure) (fig 2).

Mechanosensitive pelvic nerve sensory fibres that innervate the urinary bladder or distal colon have low (<5 mm Hg) or high (>30 mm Hg) thresholds for response to distension. Both low (LT) and high (HT) threshold sensory fibres encode the distending pressure well into the noxious range (>40 mm Hg). (Modified from Sengupta and Gebhart.3)
Visceral mechanoreceptors that have low thresholds for response (for example, <5 mm Hg) respond within the physiological range. Interestingly, as illustrated in fig 2, these afferent fibres also respond to distending pressures in the noxious range (>30 mm Hg) and encode stimulus intensity fairly well. Note also that response magnitude in the noxious range is typically greater than response magnitude of high threshold fibres at the same distending pressure. Visceral mechanoreceptors that have high thresholds for response do not begin to respond until the distending pressure is at or exceeds pressures that are likely noxious.
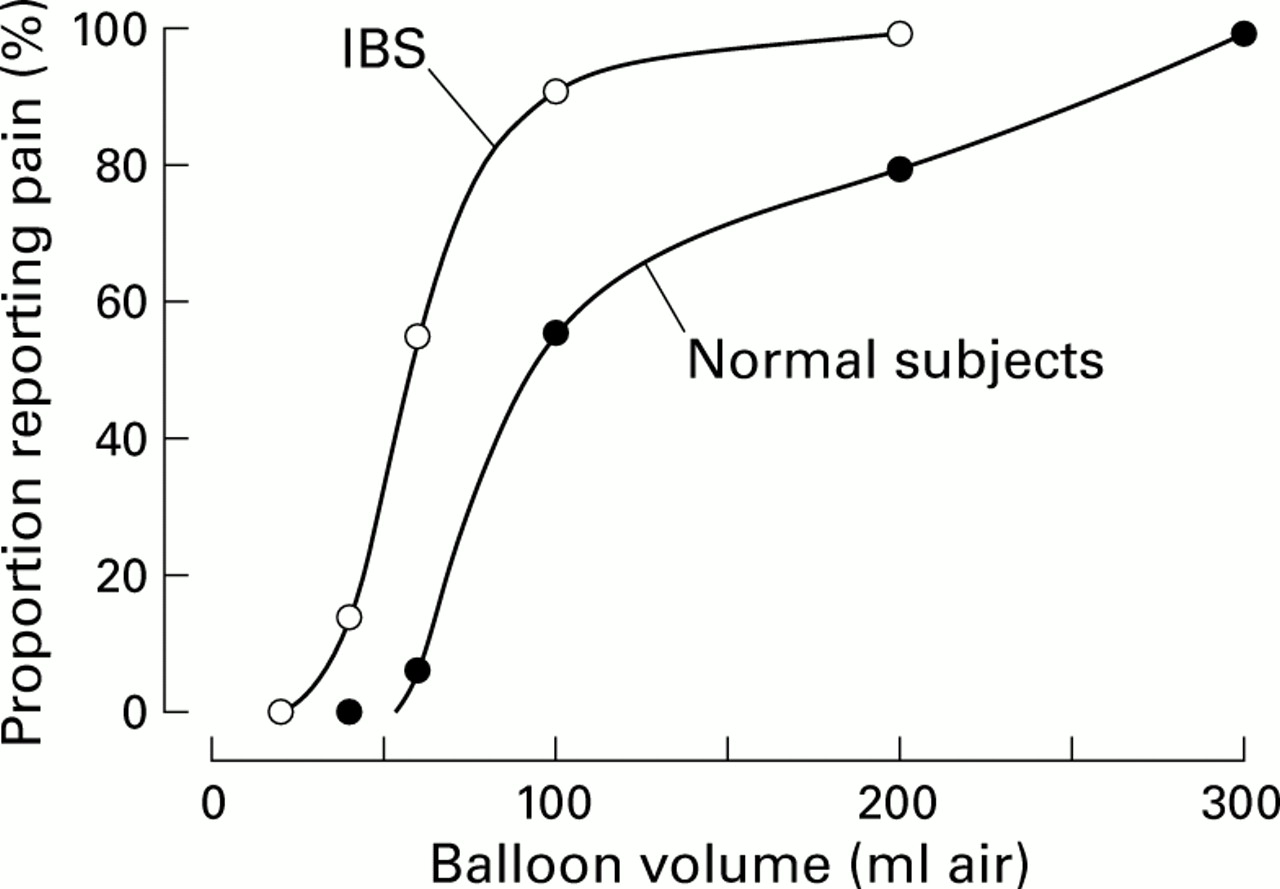
How do responses of visceral sensory fibres change in response to visceral inflammation? As they typically do, clinical observations led the way to changes in the way in which visceral pain is considered. In 1973, Ritchie4 documented that irritable bowel syndrome (IBS) patients reported pain at lower volumes of balloon distension of the colon than normal subjects (fig 3).

Illustration of the presence of visceral hyperalgesia in patients with irritable bowel syndrome (IBS). (Modified from Ritchie.5)
As illustrated, less than 10% of normal subjects reported pain at a distending volume of about 60 ml whereas greater than 50% of IBS patients reported pain at the same distending volume. This leftward shift in the psychophysical function of IBS patients suggests the presence of hyperalgesia in IBS. Qualitatively similar outcomes have been confirmed subsequently by others and also extended to other hollow viscera in patients with other of the so called functional bowel disorders (non-cardiac chest pain, non-ulcer dyspepsia, and IBS). Accordingly, it is considered that the altered sensations (bloating, discomfort, and abdominal pain) associated with these functional disorders represent visceral hyperalgesia.6 If this hypothesis is correct, then visceral afferent fibres should exhibit sensitisation and the spinal neurones on which they terminate should undergo a change in excitability, mechanisms of primary and secondary hyperalgesia, respectively (see discussion in Mayer and Gebhart6).
Experimental inflammation of hollow organs has been shown to sensitise responses of mechanosensitive visceral sensory fibres and to awaken so called silent afferent fibres. By sensitisation is meant an increase in response magnitude to distending stimuli, sometimes associated with a reduction in response threshold and/or an increase in spontaneous activity. That is, the excitability of the sensory neurone is increased, as illustrated (fig 4) for a high threshold mechanosensitive pelvic nerve fibre innervating the colon of the rat.
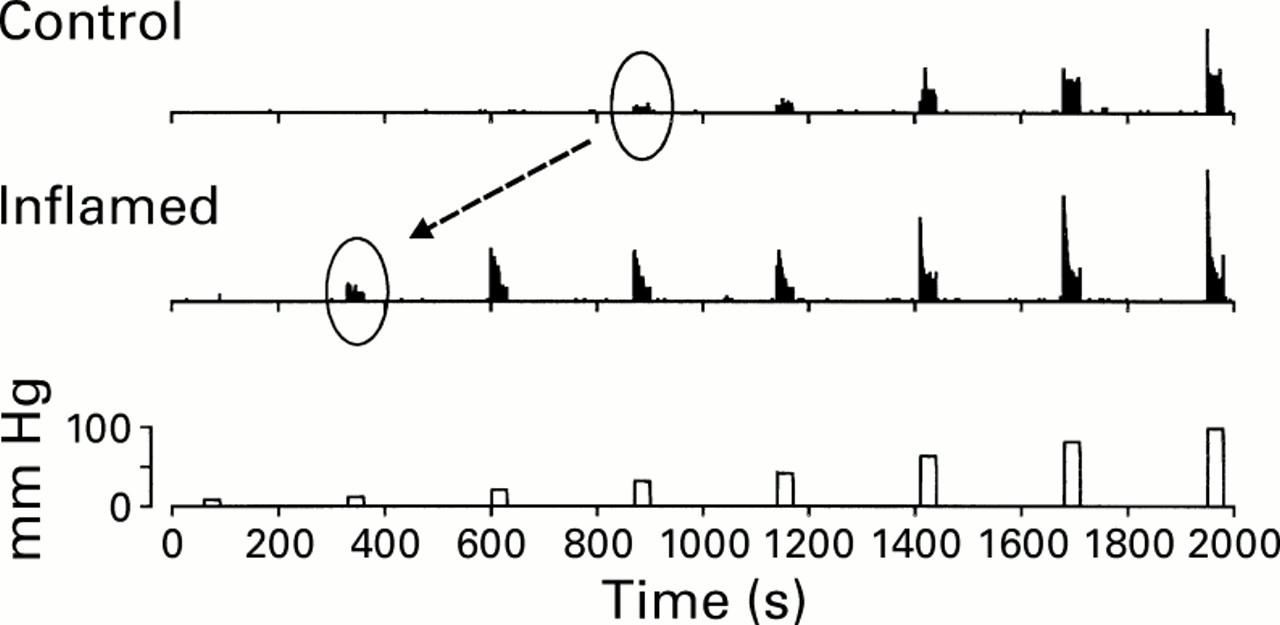
Visceral sensory fibres sensitise to experimental organ inflammation. In this example, responses of a high threshold pelvic nerve sensory fibre are shown before (control) and 30 minutes after intracolonic instillation of 2.5% acetic acid (inflamed). Note that response magnitude to distension (bottom) increases after inflammation and that response threshold decreases from about 30 to 10 mm Hg.
In this example, response threshold decreases from 30 to 10 mm Hg and response magnitude is increased for all distending pressures. This sensory fibre had no spontaneous activity and inflammation did not provoke any change in resting activity. Another consequence of visceral inflammation is the awakening of what have been termed “silent nociceptors,” which are really mechanically insensitive afferent fibres that acquire mechanosensitivity after visceral inflammation. Such silent or sleeping afferents are present in all tissues studied (including human skin), are normally unresponsive to acute noxious stimulation, and are believed to add to the afferent barrage received by the central nervous system when tissue is injured. An example of a silent afferent fibre innervating the colon of the rat is illustrated above. This fibre did not respond to either high intensity colorectal distension (CRD) or urinary bladder distension, the principal organs innervated by the pelvic nerve. After intracolonic instillation of mustard oil, a C fibre excitant, the fibre became active and mechanosensitive (fig 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Example of a mechanically insensitive (silent) pelvic nerve sensory fibre. Before intracolonic instillation of the C fibre excitant mustard oil, this fibre did not respond to 100 mm Hg colonic distension (CRD) (not shown). After mustard oil treatment, the fibre began to discharge and acquired sensitivity to CRD. (Modified from Sengupta and Gebhart.1)
Footnotes
- Abbreviations used in this paper:
- IBS
- irritable bowel syndrome
- CRD
- colorectal distension