Article Text

Statistics from Altmetric.com
The challenge of host defence in the digestive tract
When viewed from the perspective of host defence, the mammalian digestive tract presents many challenges. At the outset food contains various micro-organisms that could thrive in the rich growth medium of digested nutrients. Because a large surface area is required for the absorption of nutrients, there are abundant potential sites for microbial attachment and invasion in the long digestive tube with its many folds, villi and microvilli. The requirements of efficient nutrient absorption also place limits on the barrier components of host defence. Indeed, unlike the skin, vaginal epithelium or oral mucosa, intestinal mucosa is comprised of only a single layer of epithelial cells. This delicate barrier is further threatened by the effects of acid, bile salts, and hydrolytic enzymes required for digestion. Although some of these molecules are toxic to microbes, and hence may help to control microbial proliferation and survival, these agents may also subtly damage the mucosa and compromise its barrier functions. The potential for cumulative damage may explain why this epithelium is rapidly and continually replaced throughout the lifetime of mammals. As a result of the need for epithelial renewal, stem cell proliferation and differentiation are critical for normal function of the gastrointestinal tract. Damage to, or parasitisation of stem cells would have severe consequences for the maintenance of the normal digestive epithelium.
A continuous microbial threat is also posed by a wide array of colonising microbes throughout the gut, with especially large numbers in the mouth and colon. Infectious complications that often follow treatment with antibiotics suggest that colonising symbionts contribute to host defence against pathogens. Yet, in the absence of effective defence mechanisms, even symbiotic microbes can multiply rapidly and overwhelm the mammalian host. Despite the presence of microbes, normal digestive tract surfaces generally show little evidence of inflammation, the coordinated pattern of vasodilatation, leucocyte recruitment and transudation of leucocytes and plasma that constitutes a common response of many tissues to microbes. In the absence of inflammation, steady state defence in this organ system must rely on other broadly effective mechanisms. This review will focus on one such mechanism, the production of defensins, a group of antimicrobial peptides that contribute to innate immune defences in the gastrointestinal tract and other body surfaces.
Antimicrobial peptides
Antimicrobial peptides (arbitrarily defined as <100 amino acids in size) are found abundantly in many host defence settings, from plant seeds and arthropod haemolymph to human neutrophils and epithelia.1-5 Unlike many other antibiotics in nature, which are produced by specialised metabolic pathways, antimicrobial peptides are encoded by genes and produced by conventional transcription and translation. The various peptides are structurally diverse and range from simple, α-helical linear molecules to molecules with β-sheet conformation and multiple disulphide linkages, but in general have antimicrobial activity against a broad range of microbes at micromolar concentrations. Most of these peptides are cationic and amphipathic, features that confer affinity for microbial membranes rich in anionic phospholipids, and a degree of protection for host cell membranes that contain neutral phospholipids and cholesterol. Proposed mechanisms of microbicidal activity for the various peptides include pore formation and membrane depolarisation, disruption of bacterial energy metabolism and interference with biosynthetic pathways.
Evidence for the specific roles of antimicrobial peptides in host defence has been provided by experiments that assess the impact of ablation or augmentation of antimicrobial peptide production. Augmentation of antimicrobial peptide production in plants increases their resistance to plant pathogens,6-8 whereas ablation of the pathways that induce the production of antifungal peptide drosomycin in Drosophila dramatically reduces survival after fungal infections.9 Analogous experiments in transgenic mice are a subject of intense effort in several laboratories. The experimental design may be complicated by the multiplicity of antimicrobial peptides and redundancies in the innate and adaptive immune systems.
In mammals, defensins are one of the major families of antimicrobial peptides.5 Characteristically, these peptides are 30–42 amino acids in length, have a cationic charge, a predominance of β-sheet conformation, and contain six cysteines that participate in three intramolecular disulphide bonds. Based on the spatial distribution of the cystine linkages, the mammalian defensins may be classified into two major groups termed α- and β-defensins. In α-defensins, invariant disulphide bonds form between cysteines C1–C6, C2–C4, and C3–C5.10 In β-defensins, the pairing occurs between C1–C5, C2–C4, and C3–C6.11Although their disulphide linkages differ, the three dimensional structure of both groups of peptides is very similar (fig 1). In humans, the genes that encode α- and β-defensins are clustered in a segment a few hundred kilobases long of the short arm of chromosome 8 (8p23),13-15 and the corresponding homologous genes are similarly clustered in the rodent genome.16-20

Structure of α- and β-defensins. Figure reproduced from reference 12 with permission.
The α-defensins were among the first antimicrobial peptides to be discovered and are major constituents of the primary granules of mammalian granulocytes.21-23 In humans, the α-defensins HNP 1–4 account for roughly 5% of the total cellular protein of neutrophils. Defensin granule concentrations are probably in excess of 10 mg/ml and upon fusion of primary granules with a phagosome, these defensins are delivered to the engulfed micro-organism with little dilution.23 Other α-defensins are abundantly expressed in Paneth cells of the small intestinal crypts.24 ,25 In humans, two such Paneth cell α-defensins have been identified, HD-5 and -6. Recently, the non-haematopoietic α-defensin HD-5 has also been detected in epithelial cells of the genital tract.26Analysis of α-defensin gene structure in several species reveals that all haematopoietic α-defensin genes have three exons, but that the epithelial α-defensin genes have two.
The β-defensins were discovered in cattle as antimicrobial peptides of airway epithelial cells.27 Since then, expression of β-defensins in many other epithelial cell types and other species has been reported.28 Along the alimentary tract, β-defensins have been found in the gingival epithelia, the tongue and the colon.29-32 Two human β-defensins have been identified, HBD-1 and -2, with peptides (or transcripts) found in the kidney, skin, pancreas, gingiva, salivary gland, cornea, and airway epithelium.33-36 Similar β-defensins are also expressed in the granulocytes of some vertebrates, including cattle and other ruminants and some avian species.37 Like the epithelial α-defensins, all β-defensin genes characterised so far have two exons.28
Defensins in small intestinal Paneth cells
Initially, α-defensin genes expressed in Paneth cells of mice were detected as an abundant class of developmentally regulated mRNA.38 Human Paneth cell defensin genes were later identified by their sequence similarity to myeloid defensins.39 ,40 In mice, there is evidence for the expression of 19 or more (highly similar) defensin genes in the small intestine,41 whereas only two human defensins, HD-5 and -6 have been found at this site following extensive molecular searches.42 In mice, cryptdin-1 and cryptdin-5 mRNA content is equivalent in duodenum, jejunum, and ileum.43In contrast, cryptdin-4 mRNA is absent or present at very low levels in proximal small bowel, increasing to maximal levels in the distal ileum. In humans, HD-5 and -6 mRNA seem to be coordinately expressed in Paneth cells at a ratio of about 4:1. Mature enteric α-defensin peptides, also termed cryptdins, have been isolated from the murine small bowel.24 ,41 ,44 ,45 Similar attempts to isolate human peptides systematically from intestinal tissue and lumen are currently underway (see later). One isoform of HD-5 (64–94) was prepared recombinantly using a baculovirus/insect cell expression system46 ,47 and has been used to initiate functional studies of this peptide.
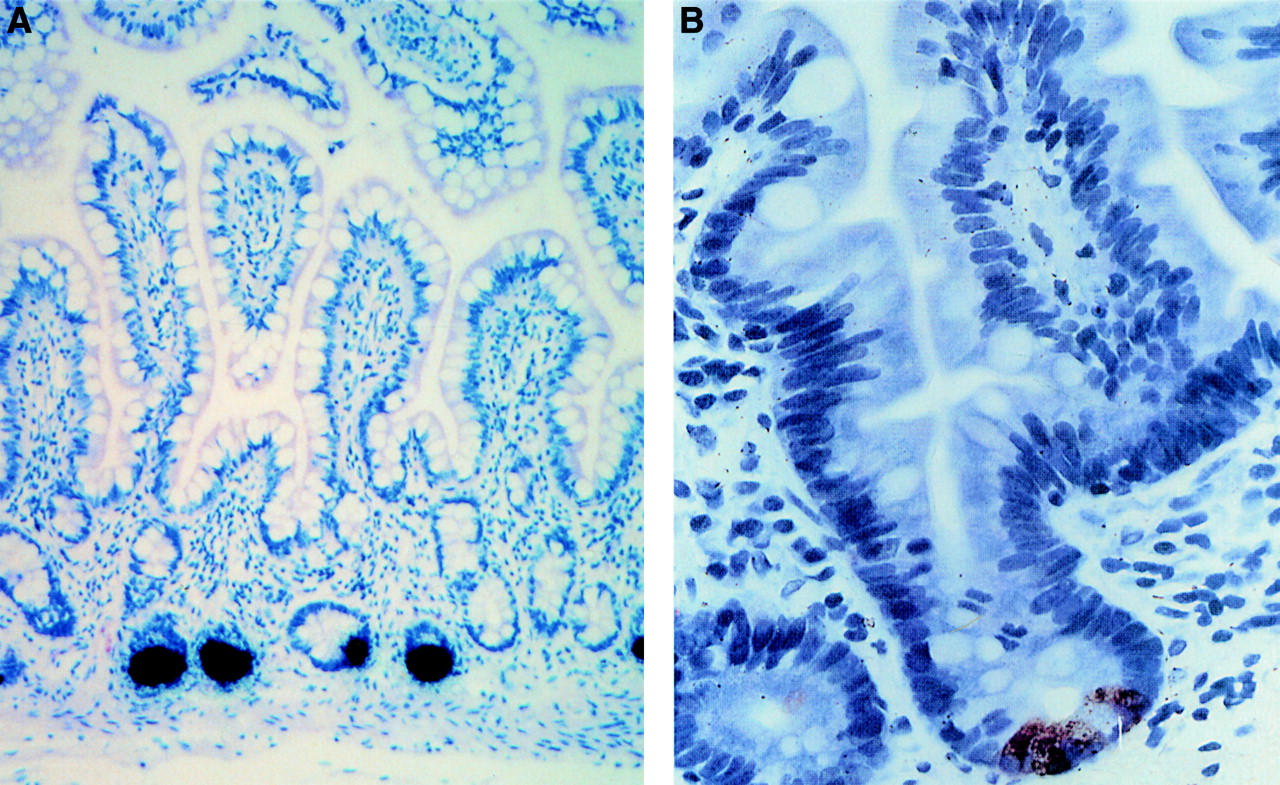
Paneth cells are granulated secretory epithelial cells residing in the bases of the crypts of Lieberkühn. Immunohistochemical studies have localised α-defensins to apical granules in these cells (fig2).24 ,46 In addition to defensins, their granules contain additional antimicrobial molecules, including lysozyme and sPLA2.48 ,49 Paneth cell granules are released to the luminal mucosal surface consistent with their proposed functions in the crypt lumen. Both intraluminal bacteria and lipopolysaccharide can stimulate this Paneth cell secretion,50 as can cholinergic agonists.48 The secretion is inhibited by atropine, a muscarinic antagonist.51 Paneth cell distribution is generally limited to the small intestine. Paneth cell hyperplasia and metaplasia can be seen in chronic inflammation associated with Barrett's oesophagus, gastritis, carcinoma, Crohn's disease, and ulcerative colitis.52 ,53
Ontologically, Paneth cell differentiation occurs during crypt morphogenesis. Subsequently, Paneth cells differentiate from stem cells that migrate downwards in the crypt.54 ,55 Tissue specific expression of Paneth cell defensins is, at least in part, transcriptionally regulated. Analysis of lines of transgenic mice carrying 6.5 kilobases of 5′-flanking sequence of the cryptdin-2 gene fused with a human growth hormone reporter gene show expression of the reporter protein in Paneth cells only.54 Transgenic mice carrying the HD-5 gene, including 1.4 kilobases of 5′-flanking sequence, express the peptide in Paneth cells in the same developmental sequence as the endogenous mouse genes (Salzman N, Bevins CL, and Huttner KM. Abstract 132/E-53 American Society of Microbiology. New Orleans, 1996). Thus, the 5′-flanking regions of the enteric α-defensin genes seem to be sufficient for direct expression in Paneth cells.
Post-translational modifications of defensins
The main post-translational modifications of defensin peptides are (a) formation of three intramolecular disulphide bonds10 ,11 and (b) proteolytic processing of the NH2-terminus to yield the mature peptide.5 ,56 In addition, in some defensin peptides the NH2-terminal glutamine is converted to a pyroglutamate residue.29 ,37
Studies of post-translational processing of defensins have focused on human neutrophil α-defensins.57-61 The human neutrophil peptides are synthesised as 93–94 amino acid prepropeptides that contain N-terminal endoplasmic reticulum targeting (signal) peptides, acidic propeptide segments of approximately 40 amino acids, and a mature peptide at the C-terminus. The defensin precursors are initially cleaved to produce intermediates of 75 and 56 amino acids and subsequently stored in granules almost entirely as fully processed mature peptides (29–30 amino acids).58 ,59 The acidic propeptide segment may be important for neutralisation, processing, and/or folding of the cationic C-terminal defensin peptide.60 ,61
The post-translational processing of epithelial α-defensins has not yet been systematically investigated, but may be a key regulatory step in the generation of active Paneth cell α-defensins. Isolation and characterisation of HD-5 peptides in human specimens have revealed an unexpected complexity in the N-terminally processed forms of this peptide.62 In initial studies of ileal tissue, at least three isoforms were detected by using western blot analysis. Two forms were characterised chemically and corresponded to amino acids 23–94 and 29–94.62 The detection of these larger forms suggests that human Paneth cells, in contrast to neutrophils, may store α-defensins as propeptides. These propeptides would then be predicted to be processed to mature peptides either during or after exocytic secretion. The study of HD-5 in a different biological context has detected shorter extracellular forms of this peptide. The surgical construction of neobladders from ileal tissue provides an opportunity to isolate Paneth cell products from voided urine. In neobladder urine, three major forms were characterised chemically, representing amino acids 36–94, 56–94, and 63–94.62 The shortest form (63–94) is similar to the mature recombinant HD-5 peptide (amino acids 64–94), which has broad spectrum antimicrobial activity.47 For epithelial β-defensins, generation of forms with variable N-termini was also observed with renal human β-defensin-1 released into voided urine.36
Defensin activities
Paneth cell α-defensins show microbicidal activity against a variety of microbes. Recombinant human Paneth cell HD-5 (64–94) is active against Listeria monocytogenes,Escherichia coli,Salmonella typhimurium, and also against the fungus Candida albicans.47 The recombinant HD-5 peptide retains activity even in presence of physiological concentrations of trypsin, evidence that this peptide could function within the hostile environment of the intestinal lumen.47 The activities of the natural isoforms of HD-5 recently identified in human small intestine tissues and from ileal neobladder urine have still to be thoroughly investigated.
More extensive structure–function studies have been possible for the mouse Paneth cell α-defensins. Of the mouse cryptdins, cryptdin-4 is the most cationic and has the greatest antibacterial activity.41 Mouse cryptdins 1, 2, 3, and 6 differ from each other in sequence only by one to four amino acids, yet these peptides have distinct in vitro activities. For example, cryptdin-2, which differs from cryptdin-3 only by a substitution of threonine at position 10 for lysine, was not bactericidal for E coli, but cryptdin-3 had potent activity. However, with respect to the sensitivity of Giardia lambliatrophozoites, cryptdins-2 and -3 were highly active, whereas cryptdins-1 and -6 had little or no effect on trophozoite survival.63 This further supports the notion that apparently subtle aspects of primary structure are important for the biological activity of the defensins. A systematic analysis of the effects of amino acid substitutions on antimicrobial activity against a variety of micro-organisms may uncover important structure–function relations that could have implications for the design of synthetic antibiotics.
In addition to antimicrobial activity, certain defensins have exerted remarkable effects on host cells.64-67 When applied to the apical surfaces of T84 cells, cryptdin-2 and -3 induce chloride secretion.68 Although not a feature of all defensins, such a secretory response induced by microbial entry into the crypt could contribute to host defence by “flushing out” the crypt. Certain other defensins also display chemotactic activity for T lymphocytes66 and other immune cells and may serve as a link between innate and adaptive immunity.69
Genetic ablation of Paneth cells
Introducing artificial transgenes consisting of a cell-type-specific promoter and a region encoding a cytotoxic protein can ablate specific cell lineages during development. This strategy has lead to valuable insights into the function of specific cell populations. Paneth cell deficiency70 was induced in mice by placing the 5′-flanking region of the mouse cryptdin-2 gene upstream of an attenuated diphtheria toxin A fragment coding sequence. In these transgenic mouse lines, roughly 95% of Paneth cells were eliminated and apparently undifferentiated crypt columnar cells occupied the Paneth cell deficient crypts. In the crypts of these mice maintained in a barrier facility there was no evidence of colonisation by commensal gut microflora as detected by silver staining.70 These data suggest that Paneth cell factors may not be required to prevent colonisation of the small intestinal crypt by commensal flora, or that the remaining 5% of the Paneth cells and other factors can keep the crypt germ-free in otherwise healthy mice. Also, in the absence Paneth cells, it is possible that the epithelium continues to express Paneth cell genes or that other antimicrobial factors are able to compensate for Paneth cell loss. It will be of great interest to test the response of the gut epithelium of these transgenic mice to pathogenic bacterial challenges.
Upregulation of HD-5 and HD-6 mRNA in necrotising enterocolitis
Necrotising enterocolitis (NEC) is a common, often devastating complication of uncertain aetiology in premature infants and is characterised by intestinal necrosis with haemorrhage and mucosal oedema.71 ,72 In cases of NEC, the levels of HD-5 and -6 mRNA expression per Paneth cell are increased threefold over that observed in controls.73 This suggests that a mediator produced in the course of NEC can induce defensin expression. In addition, the number of Paneth cells is increased about twofold in the NEC specimens compared with controls, suggesting that the disease process promotes Paneth cell hyperplasia.73 Interestingly, the intracellular concentration of HD-5 peptide as detected by immunohistochemistry seemed to be unchanged despite an increase in HD-5 mRNA in the NEC samples. One interpretation of these observations is that the defensin peptides are secreted at an increased rate in response to the ongoing disease process.
The role of defensins in intestinal host defence: a hypothesis
The location of defensin secreting Paneth cells at the bottom of a long narrow crypt adjacent to the mitotic stem cells suggests that defensins act at a high local concentration soon after secretion, and that the object of their activity is the protection of the epithelial stem/progenitor cells of the small intestine (fig 3). If Paneth cells contain as much defensin peptide as neutrophils do, the concentration of defensins at the base of the crypt could reach millimolar levels. In view of the well documented antimicrobial activity of many defensins, it seems reasonable to propose that the function of Paneth cells and their defensins is the protection of the crypt and its stem cells from microbial invasion and parasitisation. It is also possible that the more dilute secretions (perhaps in the micromolar range) that reach the intestinal lumen still contain sufficient activity to influence the resident flora. Finally, at even more remote locations from the site of secretion, very dilute (nanomolar) secretions could function as chemoattractants or provide other signals for host defence cells (lymphocytes, monocytes, dendritic cells).

{kind=link}
{kind=link}
{kind=link}
A hypothesis for the role of Paneth cell defensins in intestinal host defence. See text for details.
Future perspectives
Recent discoveries have helped focus attention on the important contributions of innate immune mechanisms in mammalian host defence. Among the mechanisms of innate immunity in the gastrointestinal tract, epithelial secretion of antimicrobial defensins seems to be particularly well developed in the Paneth cells of the crypts of Lieberkühn. The structure–function relations, biological activities and regulation of Paneth cell defensins are productive areas for further investigations. As antimicrobial peptides have been found in the stomach and the colon of other vertebrates, it is likely that as yet unidentified human antimicrobial proteins and peptides are awaiting discovery in the many epithelial cell types of the digestive tract. It will be interesting to learn how the altered expression of defensins and other antimicrobial peptides, related to genetic deficiencies, developmental immaturity, chronic inflammation or concurrent systemic disease, impacts on innate host defence. Furthermore, in addition to their antimicrobial function antimicrobial peptides may provide important links between the innate and adaptive immune systems by receptor mediated signalling to lymphocytes or other host cells. A better understanding of innate host defence mechanisms may lead to therapeutic strategies that augment the effectiveness of host defence in the digestive tract.
Acknowledgments
The authors are supported in part by grants from the National Institutes of Health AI32234, AI32738, AI40248 and HL46809.
Abbreviation used in this review
- NEC
- necrotising enterocolitis