Article Text

Statistics from Altmetric.com
SUMMARY
The peroxisome proliferator activated receptor γ (PPARγ) is a nuclear receptor highly expressed in the colon and playing a key role in bacterial induced inflammation. Regulation of colon inflammation by this receptor has been well demonstrated in many experimental models of colitis but also in patients with ulcerative colitis, characterised by impaired expression of PPARγ confined to their colon epithelial cells. Recent data showing that PPARγ was the major functional receptor mediating the common aminosalicylate activities in inflammatory bowel diseases (IBD) have also reinforced the roles of this receptor in the control of intestinal inflammation. The aims of this review are to discuss the potential roles of PPARγ in the physiopathology of IBD, as well as the emerging therapeutic strategies targeting this receptor.
INTRODUCTION
Current evidence suggests that Crohn’s disease (CD) and ulcerative colitis (UC) result from a complex interplay between genetic and environmental factors, leading to an abnormal innate and adaptive immune response of the gut directed against luminal constituents in genetically determined patients. Identification of cytoplasmic receptors of bacterial peptidoglycan, namely nucleotide oligomerisation domain (NOD)2/caspase recruitment domain (CARD)15 and NOD1/CARD4, as CD susceptibility genes reinforced the pivotal role of the interactions between enteric microbes and the intestinal immune system in the physiopathology of IBD.1–3 Furthermore, recent advances in our laboratory and others also indicate the involvement of another key receptor, PPARγ, which regulates colon inflammation. This represents a new target in the development of therapeutic molecules in IBD.
PPARγ is a nuclear receptor discovered in mammals in 1993 as an orphan receptor.4 Until recently, PPARγ was known as a receptor mainly expressed by adipose tissue and involved in the regulation of insulin resistance. PPARγ is activated by antidiabetic thiazolidinedione drugs.5 In 1998, the first studies were published reporting a potential link between this receptor and intestinal diseases, originally described in colon cancer6–8 and one year later during intestinal inflammation.9 There is now emerging interest in the roles of this receptor in the regulation of gut homeostasis. Using a computerised medical literature search of all English language articles selected from the “PubMed” online database with the keywords “peroxisome proliferator-activated receptor gamma”, “inflammatory bowel disease”, “Crohn’s disease”, “ulcerative colitis”, “colitis”, “ileitis”, and “intestinal diseases”, more than 100 articles were found that reported a role for PPARγ, mainly in colon cancer and intestinal inflammation.
After a brief presentation of PPARγ and its ligands, the aims of this review are to outline the potential roles of PPARγ in the physiopathology of IBD and highlight areas for future therapeutic strategies targeting this receptor.
PPARγ STRUCTURE, EXPRESSION, AND REGULATION
PPARγ structure and function
PPARγ belongs to the nuclear receptor family consisting of a group of approximately 50 transcription factors implicated in many different biological processes and considered as important targets in the development of new drugs.10 PPARγ is an essential nuclear receptor controlling the expression of a large number of regulatory genes in lipid metabolism and insulin sensitisation, as well as in inflammation and cell proliferation.11,12 Its activation requires heterodimerisation in the nucleus of the cells with another nuclear receptor, known as the retinoid X receptor α (RXR α) (fig 1), leading to binding of this heterodimer to specific DNA sequence elements termed peroxisome proliferator response elements (PPRE).13 It has been demonstrated that these two nuclear factors play a central role in the regulation of inflammatory signalling pathways by acting on kinases and transcription factors, such as nuclear factor κB (NFκB), c-Jun, c-Fos, and nuclear factor of activated T cell (NFAT)9,14,15 (fig 2) and inhibiting mucosal production of inflammatory cytokines (interleukin (IL)-1β and tumour necrosis factor α (TNF-α))14 and chemokines,16 proliferation of inflammatory cells,17 and expression of some adhesion molecules (fig 2).18

Peroxisome proliferator activated receptor γ (PPARγ) is a nuclear receptor which forms a heterodimer with retinoid X receptor (RXR). PPARγ may be activated by different natural and synthetic ligands allowing its heterodimerisation with RXR and binding, in the nucleus of the cell, on the peroxisome proliferator response element (PPRE). This binding regulates gene expression involved in the control of many biological processes, particularly inflammation.
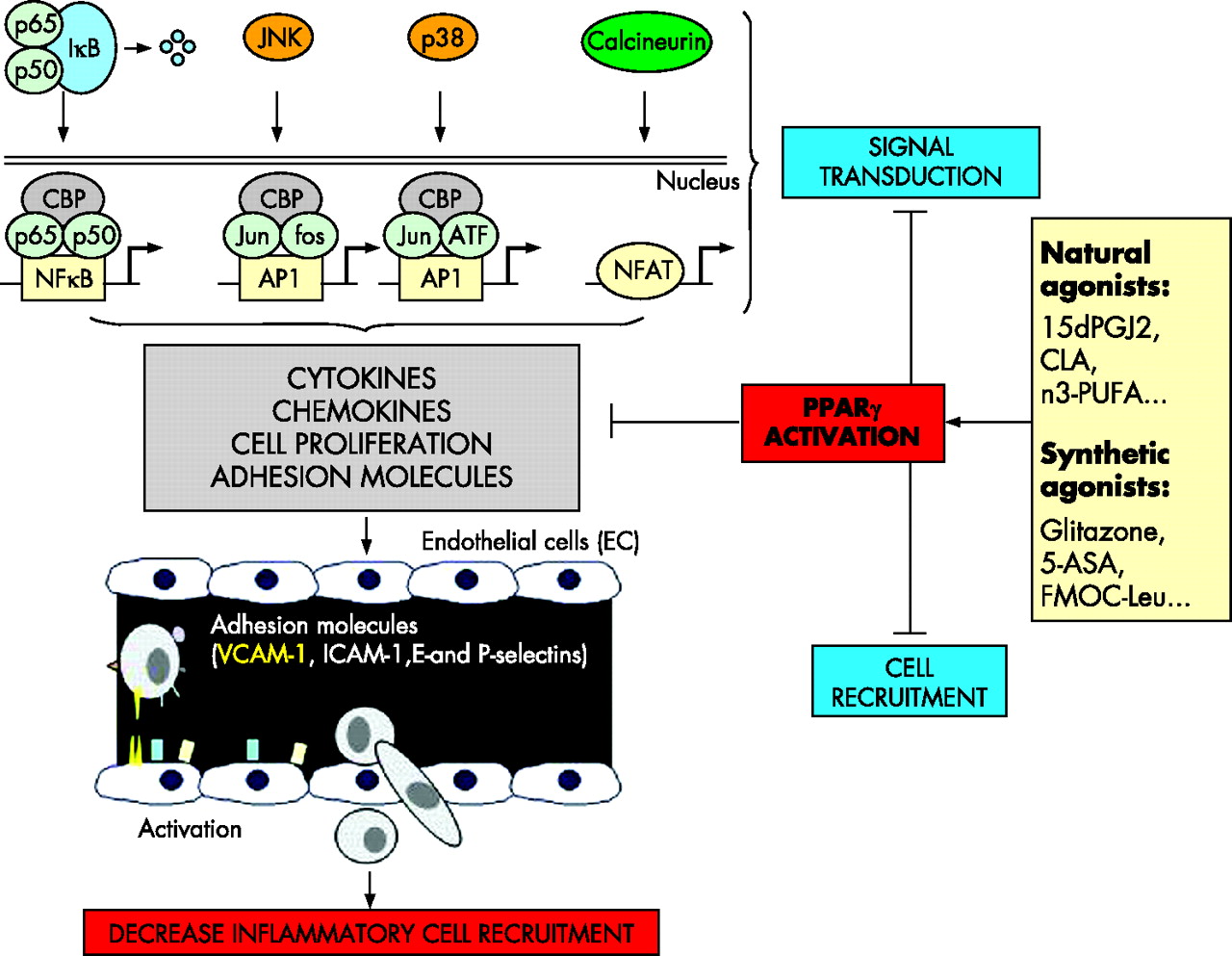
Interferences of peroxisome proliferator activated receptor γ (PPARγ) with inflammatory signalling pathways. PPARγ inhibits nuclear factor κB (NFκB) signalling pathway through interactions with NFκB, the inhibitory protein called IκB, and CBP, a coactivator of p65. The MAPK pathway is also regulated by PPARγ, which reduces JNK and p38 activation and inhibits the transcription factors c-jun, c-fos, and nuclear factor of activated T cell (NFAT). Regulation of these main signalling pathways results in inhibition of cytokine and chemokine production, cell proliferation, and adhesion molecule expression (mainly VCAM-1), which decrease inflammatory cell recruitment in inflamed tissues. 15dPGJ2, 15-deoxy-Δ12,14-prostaglandin J2; PUFAs, polyunsaturated fatty acids; 5-ASA, 5-aminosalicylic acid.
PPARγ is highly expressed in the colon
High levels of PPARγ expression have been reported in both colonic and adipose tissues. Originally described as a receptor expressed by adipose tissue where it plays a role in adipocyte differentiation and in the regulation of insulin responses, other tissues and cells are now known to express PPARγ (fig 3).19 Among them, the colon is a major tissue expressing PPARγ in epithelial cells and to a lesser degree macrophages and lymphocytes.20–24

Peroxisome proliferator activated receptor γ (PPARγ) mRNA expression in different tissues. PPARγ mRNA was quantified by reverse transcription-competitive polymerase chain reaction in different human organs and tissues. The main sources of PPARγ are adipose tissue and the colon. PBMC, peripheral blood mononuclear cells.
Microorganisms regulate PPARγ expression in the colon
PPARγ is a modestly inducible receptor. Regulation of its expression remains poorly investigated although some reports suggest that it might be dependent at least in part on the cellular environment. In vivo, PPARγ mRNA and protein levels are negatively regulated by long term hypocaloric diet,25 fasting, and insulin deficient diabetes,26 and positively by obesity and a diet rich in fatty acids.25,26 More precisely, two classical pathways acting on PPARγ expression have been commonly observed using adipocyte cell lines. Firstly, specific natural or synthetic ligands of PPARγ can induce a mean 2–3-fold expression of this receptor in a positive feedback loop.27 Secondly, different studies have demonstrated in vitro a synergistic effect of insulin and corticosteroids in inducing in vitro human PPARγ expression by cultured adipocytes.25,28 The NFκB and stress kinase pathways seem to be essential in post translational modifications of this nuclear receptor, but their regulatory effects on PPARγ expression remain uncertain. Other factors involving growth hormone,29,30 signal transducer and activator of transcription 5,31,32 and insulin growth factor 133 have also been proposed in the regulation of PPARγ expression, but these results need confirmation.29,30,34
Recent research also indicates close links between intestinal-microbial interactions and regulation of PPARγ expression by epithelial cells of the colon. To clarify the involvement of bacteria in the regulation of PPARγ expression in vivo, we showed over expression of PPARγ in the colon of mice with conventional or humanised flora compared with germ free animals.22 Similarly, in vitro studies using HT-29 and/or Caco-2 colon epithelial cells or KatoIII gastric cells have demonstrated the ability of lipopolysaccharide (LPS),22,35Saccharomyces boulardii,36 and Helicobacter pylori37 to increase by up to 2–4-fold PPARγ mRNA and protein expression. Enhancement of PPARγ expression by microorganisms is probably multifactorial and involves at least in part the LPS recognition Toll-like receptor (TLR)-4, expressed by activated epithelial cells. This was demonstrated in vivo by very weak expression of PPARγ in the colon of mice with non-functional TLR4 due to a naturally occurring mutation within the third exon of the TLR4 gene (C3H/HeJ Lpsd/Lpsdmice) compared with wild-type animals.22 These results were confirmed in vitro after transfection of Caco-2 cells with the constitutively active form of TLR4 leading to a fourfold induction of PPARγ expression (fig 4).22 An alternative way to regulate PPARγ expressed by epithelial cells through bacteria might be production of the volatile fatty acid butyrate produced by commensal intestinal flora. In contrast with other short chain fatty acids such as propionate or valerate, butyrate 2 mM caused a two- and sevenfold increase in PPARγ protein expression, respectively, after three and seven days of incubation of Caco-2 epithelial cells.38

Modulation of peroxisome proliferator activated receptor γ (PPARγ) by Toll-like receptor-4 (TLR4). Activation of TLR4 by lipopolysaccharide (LPS) induces PPARγ expression and activation in transfected Caco-2 cells.
Taken together, these results indicate the pivotal role of bacteria in the regulation of PPARγ expression by epithelial cells, which might account for the characteristic and important PPARγ pattern expression in the colon compared with other parts of the digestive tract. Although all microorganisms probably do not have the same ability to induce PPARγ expression, it seems that LPS of Gram negative bacteria are critical in colonic steady state PPARγ expression through TLR4. Studies are now in progress to evaluate the capacity of commensal bacteria to induce PPARγ expression and activation and to use this property as a criterion for probiotic selection.
NATURAL AND SYNTHETIC LIGANDS OF PPARγ
Natural ligands
Many natural endogenous lipophilic species such as the polyunsaturated fatty acids (PUFAs)39 and eicosanoids40 are classically proposed as natural PPARγ ligands (table 1). However, their intrinsically low binding affinities and weak in vivo concentrations in intestinal cells do not support physiological functions of many of these compounds.
Affinity and physiological roles of natural peroxisome proliferator activated receptor γ (PPARγ) modulators
Although many PUFA activate PPARγ in micromolar amounts and are recorded as functional in human plasma at these concentrations,39 their in vivo intestinal effects through PPARγ activation remain hypothetical as concentrations of these fatty acids within colonic cells are unknown. Recently, two studies performed by the group of Bassaganya-Riera et al demonstrated that, in contrast with a mixture of eicosapentaenoic and docohexaenoic acids, food supplemented with conjugated linoleic acid (CLA) efficiently prevents the development of colitis in pigs and mice.50,51 Moreover, they confirmed the direct involvement of PPARγ in the mechanism of action of CLA using colonic PPARγ null mice obtained by a Cre-lox recombination system.51 Chemically, CLA is a mixture of four isomers (cis-9, cis-10, trans-11, and trans-12) of linoleic acid with both distinct biological properties. As CLA is mainly found in milk and meat products and may also be generated from linoleic acid by human gut microflora,52 these studies are important, identifying for the first time that PPARγ natural ligands present in food or synthesised by commensal flora may improve colon inflammation.
The eicosanoid 15-deoxy-prostaglandin J2 (15d-PGJ2) is also proposed as a natural ligand of PPARγ.40 Preventive intravenous administration of high doses of 15d-PGJ2 (0.3 mg/kg) reduces ileal injury and mortality induced by intestinal ischaemia and reperfusion in rats.53 However, the physiological role of 15d-PGJ2 in PPARγ activation in the colon is still open for debate as minimal concentrations of 15d-PGJ2 required to activate PPARγ are approximately 10–150-fold higher that those found in human intestinal epithelial cells.54
Recently, the unsaturated fatty acid derivative nitrolinoleic acid (LNO2), generated via nitric oxide dependent oxidative inflammatory reactions, has been identified as a new PPARγ agonist.43 Present in the vascular cell wall as the most abundant bioactive oxide of nitrogen and in the blood of healthy individuals at concentrations of approximately 500 nM, LNO2 is considered at present to be one of the most potent physiological endogenous natural ligand of PPARγ. It works at nanomolar concentrations and displays 10-fold more efficacy than other known natural ligands such as lysophosphatidic acid, isomers of CLA, and 15d-PGJ2.43 If LNO2 seems interesting in vascular diseases, future studies are needed to determine its intestinal effects in the maintenance of gut homeostasis and during inflammatory disorders.
Synthetic ligands
PPARγ has a large ligand binding pocket that accommodates lipophilic ligands, belonging to several different groups of chemical compounds such as thiazolidinediones, also known as glitazones which bind selectively PPARγ, and glitazars which bind both PPARα and PPARγ. Troglitazone was the first glitazone developed for therapeutic use in patients with diabetes and withdrawn from the market due to severe hepatic toxic effects. After demonstration that liver injury of troglitazone was idiosyncratic and independent of PPARγ stimulation, numerous additional glitazone molecules have been developed and two are already approved in the treatment of type 2 diabetes (rosiglitazone-avandia and pioglitazone-actos) (table 2). Glitazar is a novel family of dual acting PPARα/γ agonists developed as an oral treatment for insulin resistance related glucose and lipid abnormalities associated with type 2 diabetes and the metabolic syndrome.55 Four glitazar molecules have been developed and are awaiting FDA approval (table 2). Non-steroidal anti-inflammatory drugs are also reported in vitro as PPARγ ligands but in vivo their binding affinities of 0.1 mM are 1000-fold higher than the mean concentrations found in patients conventionally treated with these drugs (table 2).56
Affinity and intestinal functions of synthetic peroxisome proliferator activated receptor γ (PPARγ) modulators
To date, only one open label pilot trial has evaluated the efficacy of the PPARγ ligand rosiglitazone (4 mg orally twice daily) in 15 patients with active UC, refractory to conventional treatment with either corticosteroids or immunomodulators and 5-aminosalicylic acid.69 After 12 weeks of treatment with rosiglitazone, a substantial decrease in disease activity index score was reported, with clinical and endoscopic remission (27% and 20%, respectively) or part response (27%) in eight patients.69 Due to their systemic effects, the most well known adverse events of thiazolidinediones observed in patients with diabetes are weight gain and infrequent hepatotoxicity. This study in IBD patients led to new clinical trials in IBD with these chemical compounds, and may lead to the development of safer PPARγ agonist with topical effects and targeting selectively the colon.
5-Aminosalicylic acid (5-ASA): a prototype of a new class of PPARγ agonists
Recently, we published studies showing functional, biological, pharmacological, and chemical evidence that aminosalicylates are a new functional synthetic ligand for PPARγ in colonic epithelial cells.64 5-ASA is one of the oldest anti-inflammatory agents in use for the treatment of IBD, but the mechanism underlying its intestinal effects remains unknown. We showed that chemically induced colitis in mice heterozygous at the PPARγ locus (PPARγ +/−) was refractory to 5-ASA therapy, arguing for a major role of PPARγ in mediating in vivo the anti-inflammatory effect of 5-ASA in the gut. Using the HT-29 colon epithelial cell line, we found that 5-ASA induced PPARγ expression. 5-ASA was also able to bind PPARγ, to induce its translocation from the cytosol of epithelial cells to the nucleus, to promote a PPARγ conformational change, and to recruit a coactivator named DRIP (fig 5). Docking simulations showed a binding mode of 5-ASA very similar to the crystal orientation of the thiazolidinedione head group of rosiglitazone. 5-ASA fitted tightly with the PPARγ ligand binding domain interacting via hydrogen bonding with His-323, His-449, Tyr-473, and Ser-289, considered as key determinants required for molecular recognition and PPARγ activation (fig 6).64 Taken together, these data show that PPARγ is an essential receptor mediating the common 5-ASA activities in IBD.

Molecular mechanisms of peroxisome proliferator activated receptor γ (PPARγ) activation by 5-aminosalicylic acid (5-ASA). After oral administration, 5-ASA crosses the cell membrane of the epithelial cell through a transporter and binds to PPARγ in the cytoplasm. 5-ASA then induces its nuclear translocation, promotes a PPARγ conformational change, and recruits the coactivator DRIP, leading to formation of a heterodimer between PPARγ and retinoid X receptor (RXR) and activation of the PPARγ response elements (PPRE).

Structural aspects of 5-aminosalicylic acid (5-ASA) binding to peroxisome proliferator activated receptor γ (PPARγ) ligand binding domain. 5-ASA, in green, located in the PPARγ ligand binding domain, interacts via hydrogen bonding with His-323, His-449, Tyr-473, and Ser-289, coloured by atom type, and considered as key determinants required for molecular recognition and PPARγ activation.
PPARγ IN IBD
PPARγ and experimental models of colitis
The first evidence of the involvement of PPARγ in the regulation of intestinal inflammation came from the use of the PPARγ synthetic agonist thiazolidinedione in mice with colitis induced by oral administration of dextran sodium sulfate (DSS).9 In this study, the two thiazolinediones troglitazone and rosiglitazone dramatically reduced disease severity in mice with colitis from 47% to 70%, seven days after DSS administration compared with the placebo treated group. These results were confirmed and extended several months later in another model of experimental colitis induced in mice by intrarectal administration of 2,4,6-trinitrobenzene sulfonic acid (TNBS). Thiazolidinediones given preventively or in treatment mode have a therapeutic effect, reducing mortality, intensity of macroscopic and histological lesions, and levels of biological markers of colon inflammation, including the NFκB and stress kinase pathways involved in transduction of inflammation.14 In addition, genetic involvement of PPARγ in the protection against colon inflammation was shown by the increased susceptibility of PPARγ heterozygous mice (PPARγ+/−) to TNBS induced inflammation compared with their wild-type littermates.14 At the present time, more than 20 published studies have reported similar prophylactic and therapeutic effects of PPARγ in different strains of mice, rats, or pigs with acute colitis induced by chemical compounds,9,14 bacteria,50 ischaemia-reperfusion,70 and also in chronic colitis occurring after the transfer of immunocompetent T cells in SCID mice51 or spontaneously in IL-10 deficient mice71 and SAMP1/YitFc animals (table 3).72
Anti-inflammatory properties of peroxisome proliferator activated receptor γ (PPARγ) in experimental models of inflammatory bowel diseases
Lessons from these animal studies are numerous. Firstly, natural and synthetic ligands of PPARγ are both effective in the treatment of acute and chronic colitis, with a similar beneficial effect of CLA and thiazolidinediones. Secondly, even if these treatments are efficacious when they are administered preventively or in treatment mode, a prophylactic effect is always more pronounced suggesting that PPARγ agonists may have higher efficacy in maintenance than in induction treatment in IBD patients. Thirdly, the therapeutic effect of PPARγ is mainly dependent on its abundance in target tissues. This notion is supported by the different susceptibility to colitis of animals in which the PPARγ gene has been disrupted14,51 or enhanced through gene transfer using adenoviruses,73 and also by analysis of SAMP1/YitFc animals where specific impaired expression and activation of PPARγ in the crypts of the small intestine is associated with ileitis.72 As PPARγ is expressed in the colon by epithelial cells and lamina propria mononuclear cells such as macrophages, and T and B cells, additional investigations in animals with cell type specific expression of PPARγ are required to determine the main cellular source responsible for the therapeutic effect of PPARγ.
PPARγ in patients with ulcerative colitis and Crohn’s disease
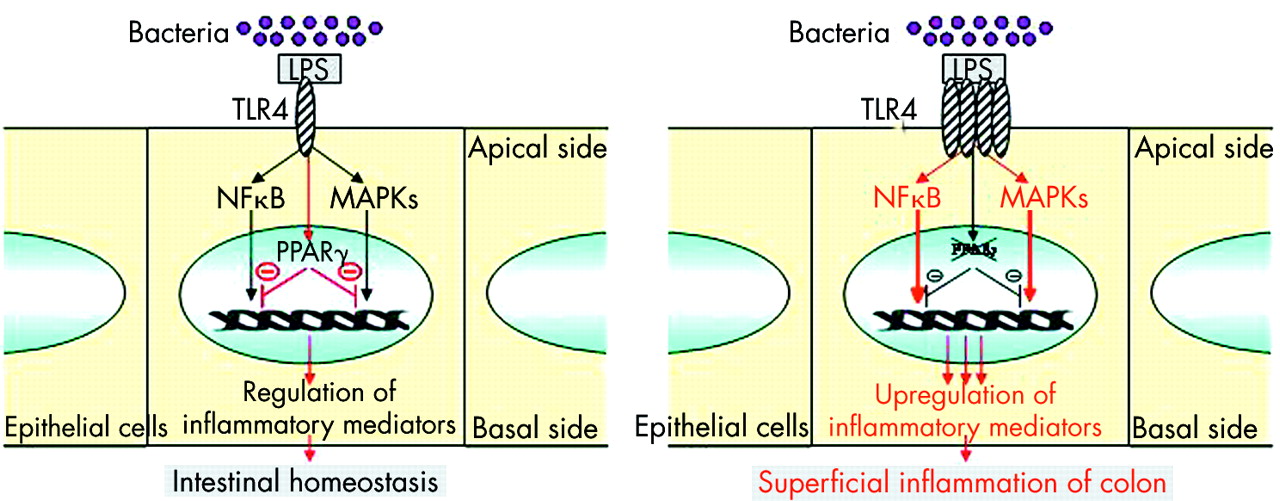
Despite in vitro and in vivo evidence of the anti-inflammatory functions of the PPARγ/RXR heterodimer in the colon, very few studies have assessed the role of PPARγ in UC and CD.22,69,72 As PPARγ is mainly expressed in the colon by epithelial cells, prior expectation might be that decreased expression of this receptor may be found in an inflammatory disorder confined to superficial layers of the intestine and limited to the colon, such as UC rather than CD. Using quantitative polymerase chain reaction, ribonuclease protection assay, western blot, and imuunohistochemical methods, 60% decreased expression of PPARγ was observed at the mRNA and protein levels in the colon of UC patients compared with patients with CD and controls.22 This impaired expression was found in both healthy and inflamed colon and was limited to epithelial cells, suggesting that perturbed levels of PPARγ in UC are not secondary to the inflammatory process. The aetiology underlying impaired PPARγ expression in colonic epithelial cells of UC patients remains unknown. Comparable levels of PPARγ in peripheral mononuclear cells of IBD patients and controls and absence of specific mutations of the PPARγ gene or its promoter in UC patients suggest that epigenetic events may account for impaired PPARγ expression in UC patients.22 Another attractive possibility may be that TLR4 signalling to PPARγ is impaired in UC and an imbalance between elevated levels of TLR422 and impaired expression of PPARγ in epithelial cells of UC patients may alter mucosal tolerance to luminal LPS, resulting in superficial colonic inflammation (fig 7). More generally, we can hypothesise that impaired expression of PPARγ in UC may be secondary to non-functional regulation of PPARγ expression in epithelial cells due to abnormal signalling pathways and/or lack of luminal stimuli induced by natural ligands or microorganisms. Further study is required to investigate more precisely the complex regulation of PPARγ expression by epithelial cells in UC patients.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Physiopathological model integrating impairment of peroxisome proliferator activated receptor γ (PPARγ) regulation by Toll-like receptor 4 (TLR4) in patients with ulcerative colitis.19 Induction of PPARγ expression in intestinal epithelial cells by lipopolysaccharide (LPS) activated TLR4 leads to regulation of nuclear factor κB (NFκB) and MAPK pathways and control of the inflammatory response. Upregulation of TLR4 expression together with impaired expression of PPARγ in epithelial cells may lead to superficial colonic inflammation in patients with ulcerative colitis.
More recent data suggest that the role of PPARγ in the physiopathology of IBD will not be limited solely to UC but may also involve CD. Based on SAMP1/YitFc animal findings developing spontaneous ileitis due to a defect in expression of PPARγ in ileal crypts, secondary to inheritance of AKR alleles in the region of PPARγ, Sugawara et al tested the relationship between PPARγ alleles and CD in humans. They demonstrated that two intronic polymorphisms SNP1 (p<10−5) and SNP2 (p⩽10−3) exhibited lower allele frequencies in 134 CD patients compared with 125 controls.72 Replication of these results in independent cohorts of patients, family based analyses, and genotype/phenotype correlation studies will be necessary to conclude more definitely that PPARγ is a susceptibility gene in CD.
CONCLUSION AND PERSPECTIVES
PPARγ is highly expressed in the colon and a key receptor in the regulation of intestinal inflammation induced by bacteria. Other studies also indicate a role of PPARγ in tumour suppression, particularly in colon cancer.6–8 Therefore, greater knowledge of PPARγ expression and function in intestinal homeostasis and during inflammation will fuel speculations about its potential therapeutic effects in IBD to prevent inflammation and colorectal cancer.81 The discovery that 5-ASA is a new topical ligand for this receptor expressed by colonic epithelial cells paves the way for the development of new molecules specifically targeting intestinal PPARγ. Because 5-ASA was originally developed without any prior knowledge of its molecular target, there is hope that the research described above will lead to rationale optimisation or development of better PPARγ ligands. To date, 20 new molecules have been developed, optimised by docking analysis to activate PPARγ in intestinal epithelial cells. Among them, two families of compounds have been selected having 30–50-fold more efficacy than 5-ASA in activating PPARγ (personal communication). Optimisation of these new molecules is now in progress. Improvements in efficacy and safety may reside not solely in new compounds with higher affinity but also in a combination of agents with additive or synergic effects on PPARγ/RXR heterodimer. In this way, studies showing the synergistic effects of PPARγ and RXR agonists must be considered.14 Furthermore, of considerable interest is the recent discovery that some commensal bacteria and natural ligands present in food may induce PPARγ expression and activation in the colon. These data suggest the potential of associating a natural regulator and a synthetic ligand of PPARγ as drug therapy for IBD patients.
Acknowledgments
The authors would like to thank the Association François Aupetit, IRMAD, UCB Pharma, and Giuliani Spa for their support. We thank Dr LJ Egan for critical review of the manuscript and improvement of language style.
REFERENCES
Footnotes
-
Conflict of interest: None declared.