Article Text

Statistics from Altmetric.com
The 1990s were heralded as the “decade of the brain” amid excitement that major advances in imaging technology coupled with huge financial investment would finally allow neuroscientists to unravel the mysteries of the human brain. Several gastrointestinal disease research groups, frustrated by the lack of objective methods to assess gastrointestinal sensory dysfunction in functional gastrointestinal disorders (FGD), recognised this period as an opportunity to utilise these new brain imaging techniques and gain a novel insight into brain-gut interactions. What followed was a period of intense activity and a flurry of publications describing the cortical representation of visceral sensation appeared (for review see Derbyshire1). It seemed only a matter of time before central mechanisms for chronic gastrointestinal pain in FGD patients would be identified, and appropriate treatment strategies designed.
Nearly a decade on from the initiation of these studies, we are still waiting. The apparent failure of brain imaging to have an impact on the diagnosis and treatment of FGD has, rather like the emperors new clothes, led an increasing number of sceptics to wonder whether this has been another false dawn in the search for a solution to the problem of chronic pain in FGD. Despite such scepticism, we believe that brain imaging has greatly increased our understanding of the neuroanatomy and functional relevance of the cortical and subcortical structures involved in processing gastrointestinal sensation. The aim of this article is to guide the reader through a summary of this information, paying particular attention to its relevance to FGD.
BRAIN IMAGING: TECHNIQUES
The two main brain imaging techniques discussed in this article are functional magnetic resonance imaging (fMRI) and positron emission tomography (PET). PET relies on the principle of detecting positron emitting radionuclides that are intravenously injected into a subject positioned so that the head is placed in a specialised detection system. There are currently hundreds of radiolabelled compounds used in PET, each with their own characteristics and used for imaging particular biochemical or physiological processes.2
While PET radiopharmaceuticals can be used to mark metabolic processes such as glucose uptake into active neural tissue and labelling of molecules that bind to specific receptors in the brain, the most commonly used in gastrointestinal research is 15O, which is prepared as H215O labelled water. This approach is used to create perfusion images that measure blood flow to the brain. Cerebral blood flow has been used a surrogate measure of brain activation as increases in regional cerebral blood flow (rCBF) have been closely correlated with regional glucose consumption3 and more latterly with neuronal activity.4 These studies have predominantly utilised block design paradigms that compare images taken during active task periods and rest periods. This type of paradigm forms the principle for the majority of functional brain imaging studies discussed in this article.
fMRI produces images of active brain regions by detecting indirect effects of neural activity on local blood flow, volume, and oxygen saturation, giving detailed information regarding the functional neuroanatomy of the brain. The first fMRI images were produced using contrast agents to demonstrate an increase in blood volume in the visual cortex in response to a visual stimulus5 but it soon became clear that naturally occurring differences in the magnetic properties of blood constituents could be utilised in order to produce these images. The most commonly used fMRI technique relies on the detection of deoxygenated haemoglobin, due to its paramagnetic properties within the blood, and to identify changes in its regional distribution during conditions which change neuronal activation.6
THE PAIN NEUROMATRIX
The predominant reason why patients with FGD seek medical attention is chronic episodic abdominal pain.7 Recently, Craig has described pain as a “homeostatic emotion” unique from our conventional senses in that it is multifaceted. He describes pain as a “discriminative sensation, an affective motivation, a potent autonomic drive, and a reflexive motor stimulus”.8 This complexity was reflected in early PET studies of somatic pain9,10 which revealed activation of an intricate network of cortical and subcortical structures following painful thermal stimulation of the skin (fig 1). Therefore, understanding the neuroanatomy of gastrointestinal pain was the priority in early brain imaging studies.

Schematic diagram of the ascending pathways, subcortical structures, and cerebral cortical structures involved in processing pain. PAG, periaqueductal gray; PB, parabrachial nucleus of the dorsolateral pons; VMpo, ventromedial part of the posterior nuclear complex; MDvc, ventrocaudal part of the medial dorsal nucleus; VPL, ventroposterior lateral nucleus; ACC, anterior cingulate cortex; PCC, posterior cingulate cortex; HT, hypothalamus; S1, S2, first and second somatosensory cortical areas, respectively; PPC, posterior parietal complex; SMA, supplementary motor area; AMYG, amygdala; PF, prefrontal cortex; M1, motor cortex. All of these regions have been activated in studies of gastrointestinal pain. Adapted with permission from Price and colleagues.38
One of the first studies to describe the pain neuromatrix activated by gastrointestinal stimulation utilised PET to compare cortical activation patterns to non-painful and painful oesophageal stimuli.11 This was a relatively simple block design paradigm in which subtraction analysis was used to compare three conditions: no sensation, non-painful sensation, and pain. The resultant images (fig 2) demonstrated that the cortical regions involved in processing gastrointestinal pain were closely similar to those previously reported in studies of somatic pain. In brief, non-painful oesophageal stimulation elicited bilateral activation of the primary/secondary somatosensory cortex and insula. As would be expected, painful oesophageal stimulation increased activity within these regions but additionally activated the anterior cingulate cortex.11 Many other studies have since confirmed that these four cortical regions are the most reproducibly activated during both somatic and gastrointestinal pain paradigms.1,11–17 While it is beyond the scope of this article to discuss the functional relevance of these cortical regions to the diverse aspects of pain processing in intimate detail (for reviews see Peyron and colleagues,15 Schnitzler and Ploner,18 Willis and Westlund,19 and Craig20) it is important at this point that we provide a brief overview to help interpret the findings of subsequent brain imaging studies.

Group mean positron emission tomography data showing z score maps for the non-painful sensation-baseline (top panel), pain-baseline (middle panel), and pain-non-painful sensation (bottom panel) contrasts. Bilateral activation of the insular and primary somatosensory and motor cortices can be seen separately in both images depicted in the top panel (non-painful) whereas stronger activation of the same areas can be seen in the middle panel (painful). Furthermore, activation of the anterior cingulate gyrus is also observed in the second image of the middle panel. The main areas of interest in the scans showing the pain definite sensation contrast are the right anterior insular cortex and the anterior cingulate gyrus, which can best be seen in the bottom panel. Adapted with permission from Aziz and colleagues.11
The main function of the primary somatosensory cortex (S1) is to process tactile information but it does receive sparse nociceptive innervation.18,19 While the arrangement and characteristics of S1 nociceptive neurones points to a role in the sensory discriminatory aspects of pain processing, several studies have also shown increases in S1 rCBF when attention is directed towards the stimulus.21 This may help to explain the psychophysical findings of increased accuracy of pain localisation during directed attention.
The secondary somatosensory cortex (S2) and insula are the most commonly activated regions described in the pain literature.15,22–26 Activation in these regions is bilateral and encompasses a region comprising the depth of the sylvian fissure and the parietal/frontal operculi. While difficult to discriminate when using group imaging, due to intraindividual variability, most studies depict two distinct loci within this region corresponding to the anterior insula and posterior insula/S2. These regions encode the intensity of stimulation as rCBF increases when the stimulus intensity is increased.25
Studies which have used intracortical depth electrodes in patients with intractable temporal lobe epilepsy have revealed evidence for the functional segregation of this region. In an interesting series of experiments, electrodes were used to stimulate throughout the insula and showed that the posterior/mid insula region is part of a somesthetic network involved in processing painful and non-painful somatic sensations.27 In contrast, stimulation of the anterior insula in the same subjects elicited viscerosensitive and visceromotor responses, indicating that the anterior insula is part of a visceral network.27 Additionally, brain imaging studies have shown that the anterior insula is modulated by changes in emotional state28,29 and is strongly activated by thermal somatic stimulation.30 Posterior insula/S2 does not appear to be modulated by attention or emotion and its role appears to be in the recognition, learning, and memory of painful events.18
The anterior cingulate cortex (ACC) is also commonly activated during experimental pain research with the most frequently described activity occurring in the mid, rostral, and perigenual cingulate regions.15,31 While not involved in spatial discrimination of pain stimuli, recent studies have shown that the cingulate cortex is functionally heterogeneous, containing regions that process information regarding stimulation intensity, emotion, attention, and mood.15,32
Despite apparent similarities in the cortical representation of somatic and gastrointestinal pain, studies have revealed several subtle differences. As gastrointestinal sensation has several unique psychophysical properties,33 researchers have sought to examine the relationship between these properties and the observed differences in cortical activation patterns. Firstly, sensation arising from the gastrointestinal tract is diffuse and hard to localise when compared with that arising from somatic structure such as the skin,33 and researchers have speculated that this may be due to differential activity within the S1.
Activation of S1 has been an inconsistent finding in studies of both somatic and gastrointestinal pain. In a meta-analysis of somatic pain studies, Peyron et al demonstrated that activation of S1 was seen more consistently if phasic rather than ramp stimuli were used. In addition, the incidence of S1 activation increased when the surface area of the stimulated region was increased.15 These factors are consistent with both the sparse nociceptive afferent input to S1 and the response characteristics of nociceptive neurones therein. While it is true to say that several gastrointestinal brain imaging studies have failed to demonstrate S1 activation, Derbyshire’s recent meta-analysis of gastrointestinal brain imaging studies has shown that S1 does play a role in gastrointestinal sensory processing.1
Brain imaging studies have shown that stimulation of a limb will result in discrete activation of the appropriate homuncular region of S1, contralateral to the site of stimulus.34 In contrast, visceral stimulation elicits bilateral activation of the inferior aspects of S1.13,14 This bilateral activation is consistent with the fact that visceral organs develop embryologically from midline structures.
A more recent study provided further evidence to associate the diffuse nature of gastrointestinal sensation with its representation in S1. In this study, cortical activation patterns were compared following oesophageal distension and thermal stimulation of the anterior chest wall and demonstrated S1 activation in the homuncular representation of the trunk in response to both stimuli but oesophageal stimulation alone also activated the more inferior aspects of S1. It was concluded that the dual representation of the oesophagus in S1 was consistent with the fact that visceral pain can be referred to the skin but not vice versa.16 Taken together, these findings suggest that the diffuse nature of gastrointestinal sensation is likely to be attributed to the wide distribution of visceral and viscerosomatic neurones projecting to S1.
Another unique characteristic of gastrointestinal sensation is that it evokes strong emotional reactions.33 For instance, psychophysical studies have shown that despite matching a visceral and cutaneous stimulus for perceived intensity, the visceral stimulus is felt more unpleasantly.35 Unpleasantness can be regarded as an emotional response, mediated via limbic structures such as the ACC, and brain imaging studies have indeed shown that the ACC is either more intensely activated or different regions are activated when stimulating the visceral regions in comparison with the somatic regions of the gastrointestinal tract.13,14
Furthermore, Strigo et al reported greater activation of the ACC in response to oesophageal pain, when compared with thermal stimulation of the anterior chest wall, in a region close to the perigenual cingulate, which is thought to play an important role in our emotional response to pain.16 As this region of the ACC has dense reciprocal projections to and from a wide variety of cortical and subcortical regions, such as the hypothalamus and anterior insula,36 this implicates its role in the mediation of autonomic changes characteristically observed in response to gastrointestinal pain. So once again it was possible to relate differences in the cortical representation of gastrointestinal and somatic pain to their known physiological characteristics.
A caveat to bear in mind when interpreting data obtained when comparing two different types of stimulation is that many brain regions, such as the anterior insula and ACC, are functionally heterogeneous (that is, apparently spatially coincident regions of the cortex respond to multiple aspects of the pain experience). As metabolic imaging techniques do not have sufficient temporal resolution to differentiate between primary and secondary pain processing in these areas, accurate psychophysiological measures should be taken from subjects prior to and during brain imaging studies to ensure the validity of the conclusions derived from subsequent observations.
This brief overview of the pain neuromatrix demonstrates that brain imaging has helped us to better understand the neuroanatomy and physiology of gastrointestinal pain processing. However, activation has also been demonstrated in the prefrontal cortex, cerebellum, posterior parietal cortex, motor cortex, supplementary motor cortex, posterior cingulate, thalamus, and brainstem,1,37 and understanding the functional significance of these regions to the gastrointestinal pain experience requires further study. One way of achieving this is to examine the effects of modulating psychological and environmental factors in carefully designed experimental paradigms. The following section describes several studies that have examined such effects.
BRAIN IMAGING: CHANGING OUR PERCEPTIONS
The affective dimension of pain combines the degree of unpleasantness perceived with the emotions associated with its appraisal and future implications.38 It has long been recognised that cognitive modulation of pain can have dramatic effects on its perception.15 In FGD, a high incidence (50–80%) of psychological disorders such as heightened anxiety, depression, somatisation, dysthymia, and panic disorders have been reported.7 Therefore, a natural progression for gastrointestinal brain imaging was to examine the effects that different psychological states have on the distribution and intensity of cortical activation. Adopting this approach would greatly help in the interpretation of aberrant cortical activation patterns in future FGD studies.
Recently, the effect of negative emotional context on the brain processing of gastrointestinal sensation has been examined. Psychologists have long noted that facial expressions can provide the emotional context to our environment in which we receive information. For instance, information received while observing a facial expression of fear would associate it with a negative emotional context while a smiling face would produce an association with a positive emotional context.28,39 Therefore, this study examined the effects of negative and neutral emotional states on the processing and perception of non-painful oesophageal distension.29
The study revealed that oesophageal distension presented during a negative emotional context was perceived more intensely, induced greater anxiety, and was associated with increased cortical activity within the anterior insula and dorsal ACC when compared with distension presented during a neutral emotional context.29 These findings provided additional evidence to support the fact that changes in the pattern of cortical activity could be directly associated with psychophysiological changes, further validating the use of brain imaging to increase our understanding of gastrointestinal pain processing. More importantly, as many FGD patients are anxious, depressed, and associate gastrointestinal sensation with negative feelings,40 these findings provide evidence that such factors can alter the way we perceive gastrointestinal sensation.
The role that attentional state plays in modifying pain is the psychological variable that has been most commonly studied. Experiments have shown that pain is perceived as less intense when we are distracted from it and more intense when we focus our attention upon it. In FGD it has been shown that patients often selectively attend to sensations that arise from the gut and it has been shown that this is an important factor in sustaining symptoms.40
As gastrointestinal sensations rarely pierce our consciousness in health, it may be expected that when they do arise, processing of this sensory information may demand increased attentional resources when compared with sensory input from other modalities. This was demonstrated in a recent study which compared cortical activation patterns in response to a non-painful oesophageal stimulus with those that occurred following a visual stimulus. Selectively focusing attention on oesophageal stimuli activated regions of the brain involved in processing sensory information (S1 and S2) as well as cognitive and emotional regions (ACC and insula). In contrast, activation related to visual stimuli was restricted to the visual cortex.41
Interestingly, when selectively attending to a visual stimulus, the cortical response to oesophageal distension was significantly attenuated.41 This implies that if we focus attention on gastrointestinal sensations, as often occurs in FGD, additional neural resources are allocated to processing of this information. This may lead to an amplification of innocuous gastrointestinal sensory events resulting in increased awareness and exaggerated emotional responsiveness. It is conceivable that such a process occurs in some FGD patients and may partly explain the heightened sensory responsiveness observed in response to experimental stimuli in many FGD studies.40
The role of anticipation in pain experiments has also been systematically addressed. It has been shown that some of the cortical regions implicated in pain processing may in fact be neural correlates related to the anticipation of an imminently painful event. These regions, including the medial anterior frontal cortex, anterior insula, and posterior cerebellum, are in close proximity to regions that are activated by pain itself. It has been suggested that this arrangement may form part of a local network which helps us learn to predict impending painful events.42
A recent study which used a pavlovian classical conditioning paradigm to examine the effects of such factors on gastrointestinal sensory processing has revealed that the majority of cortical regions activated by gastrointestinal stimulation are also activated by the anticipation of such a stimulus. In this experiment, subjects learnt to associate a visual cue with a painful oesophageal stimulus. After this initial learning phase, subjects were presented with the visual cue but the oesophageal stimulation did not occur. Despite the lack of stimulation, activation was seen in S1, S2, ACC, insula, and prefrontal cortex. As the subject began to realise that the visual cue was no longer linked to oesophageal pain (extinction phase), there was a linear decrease in activity within these regions.43
This has important connotations for the design of experimental paradigms which utilise block design and employ subtraction techniques. The fact that activation of the gastrointestinal pain neuromatrix can occur in the absence of a stimulus means that it is imperative that such factors are taken into consideration when designing the all important control/baseline condition. Failure to do so, particularly when studying an anxious patient population, may lead to erroneous interpretation of results, as will be discussed in subsequent sections.
BRAIN IMAGING: DOES SEX MATTER
The higher prevalence of many FGD subtypes in the female population led researchers to explore the possibility that this may be attributed to the manner in which the different sexes process gastrointestinal sensations within the cortex. Studies of the brain processing of somatic pain have shown differential patterns of activation,44 greater activation in women,45 and greater activation in men.46 Studies of the brain processing of gastrointestinal pain have faired little better.
Kern et al examined sex differences in healthy subjects and revealed that male subjects had virtually no activity in the insula and ACC while females strongly activated these regions.47 The implication of these findings suggested that females may have enhanced sensitivity in brain regions that process the cognitive/emotional aspects of gastrointestinal sensation. However, several other studies have shown that males also activate these regions during gastrointestinal stimulation37 and no satisfactory explanation could be given for the discrepancies described between these studies.
Berman et al studied male and female patients with irritable bowel syndrome (IBS) and revealed that the insula cortex was activated more intensely in males.48 More recently, Naliboff et al have shown greater activation in the right ACC, amygdala, and ventromedial prefrontal cortex in female IBS patients when compared with males, while males have increased activation of the insula, dorsolateral prefrontal cortex, dorsal pons, and periaqueductal gray (PAG) region.49 Interpretation of the data presented by all of these studies is somewhat difficult because the normal variability and reproducibility of the brain processing of gastrointestinal sensation has yet to be adequately addressed. A recent meta-analysis of functional brain imaging data suggested relatively good concordance between studies, despite the use of different experimental paradigms and subject populations.1 However, another study which compared cortical activation in response to oesophageal stimulation in the same subjects on three different occasions suggests that the pattern of activation alters as the subject becomes accustomed to the experiment.50 This seems particularly marked in the ACC where activity becomes less intense, along with a reduction in subjective pain ratings, when the first study is compared with the second and third.50
If this variability in brain activation patterns exists in health, one can only imagine the variability to be encountered in FGD, where patients are selected on the basis of a symptom criterion rather than by specific pathophysiological mechanisms. The following section provides a brief overview of brain imaging findings in FGD and outlines some of the technical and philosophical problems encountered by researchers in this field.
BRAIN IMAGING: WHAT HAS IT TOLD US ABOUT FGD?
The previous sections have attempted to provide a concise overview of the most salient findings provided by brain imaging studies of gastrointestinal sensation. These data have not been presented in chronological order but in the order which seems most logical. However, even when presented in this fashion, it can be seen that while our understanding of normal gastrointestinal sensory processing and its modulation has greatly improved, we are still a long way from fully appreciating the diversity and subjective variability of gastrointestinal pain processing. Without sufficient knowledge of how the gastrointestinal pain neuromatrix may vary in health, and given the modulatory influences of psychological factors on this neuromatrix, it is important that data from studies in FGD patients are interpreted with caution. Otherwise, we may find ourselves resembling fortune tellers, interpreting our destiny via the pattern of tea leaves in the bottom of a cup, when trying to explain the significance of differences in cortical activity between study populations.
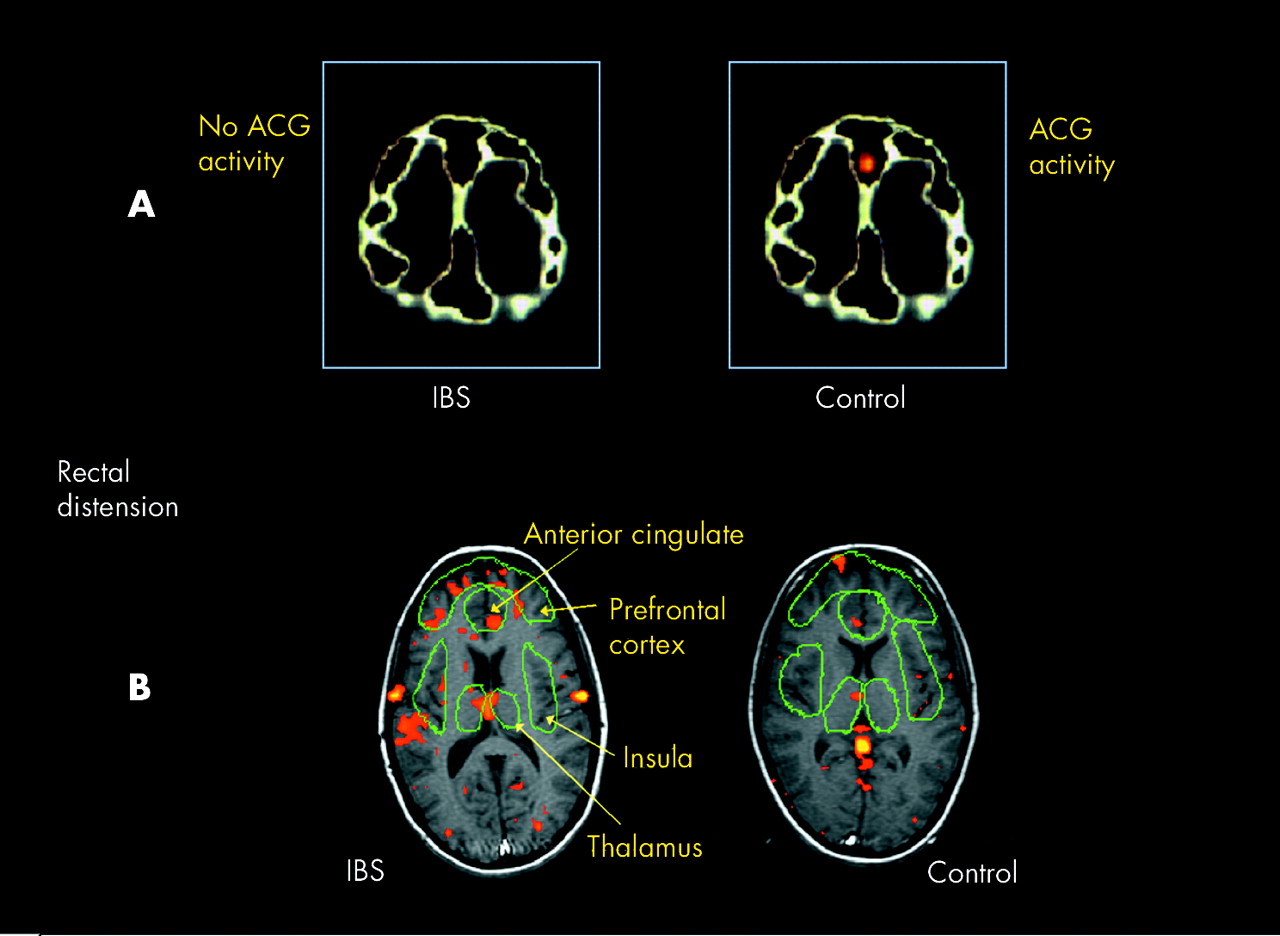
The first comparison of gastrointestinal related cortical activity between patients and healthy subjects used PET and suggested that activity in the ACC was reduced in IBS but activity in the prefrontal cortex was enhanced.51 The explanation given for these findings was that decreased ACC activity in patients may relate to a deficit in the pain modulatory effects of the endogenous opioid system while increased activity in the prefrontal cortex may relate to increased vigilance towards gastrointestinal stimulation (fig 3A). This certainly seemed to be a plausible explanation but an fMRI study several years later provided different results. In this study, IBS patients demonstrated increased activation of a different subregion of the ACC when compared with healthy subjects, leading this group to suggest that there was a heightened sensitivity of the brain-gut axis (fig 3B).52 Subsequently, researchers have reported both increased52–54 and decreased51,55 cortical activity in patients with IBS, and here in lies the problem.

(A) Images showing a comparison of cortical activity in response to rectal stimulation in a group of healthy subjects and patients with irritable bowel syndrome (IBS). It can be seen that greater activation of the anterior cingulate cortex (ACG) is observed in healthy subjects when compared with IBS. Adapted with permission from Silverman and colleagues.51 (B) Images showing a comparison of cortical activity in response to rectal stimulation in another group of healthy subjects and patients with IBS. It can be seen that greater activation of the ACG is observed this time in IBS patients when compared with healthy subjects. Adapted with permission from Mertz and colleagues.52
The aim of these studies were to identify specific pathophysiological mechanisms of visceral hypersensitivity in FGD. Two of the main hypothesise proposed as potential mechanisms of visceral hypersensitivity in FGD are:
-
gastrointestinal sensory pathways have been sensitised by a previous inflammatory or infectious event56,57 and
-
patients selectively attend to gastrointestinal sensations causing them to be amplified.40,57
Now we can ask the question, how can brain imaging help us to differentiate between these two aberrant mechanisms?
If gastrointestinal sensory pathways have been sensitised, afferent transmission of innocuous stimuli would be enhanced so that for a given stimulus intensity it would be expected that the signal arriving at the cortex would be greater in patients when compared with healthy subjects. Subsequently, activation of all brain regions involved in processing this information would be enhanced, as described in the studies by Mertz and Verne.52,53 However, several studies have shown that increasing attention towards a stimulus increases cortical activity within the thalamus, S1, S2, ACC, and PAG, in fact all of the regions described in these studies.41,58,59 Therefore, we have a problem: both proposed hypotheses fit the resultant patterns of activation so we are no closer to identifying the anomalous mechanism.
We could consider a second scenario, one in which decreased activation is observed in the mid-ACC in FGD patients. This region has been shown to be important in the cognitive processing of pain, in particular with regard to sustained attention. A patient in whom selective attention is the predominant aberrant mechanism may well be expected to have enhanced activation of this region. However, an anxious claustrophobic patient placed inside a noisy magnetic resonance scanner, with a large balloon about to be painfully inflated in their rectum, may be forgiven for sustaining their attention throughout the entire study period. In this situation, the mid-ACC may be active throughout the study and therefore subtraction analysis may reveal no differences between the two states (that is, decreased activation). Conversely, a patient with sensitised gastrointestinal sensory pathways may well show enhanced activation of this region. Once again brain imaging does not help us to differentiate between these two mechanisms. So the question remains, how can brain imaging improve our understanding of FGD?
Simply put, we need to use the right tool for the right job. Brain imaging will not answer all of the questions but used in combination with other physiological, neurophysiological, and psychological techniques it will remain an important tool in the gastrointestinal researchers armoury. The following section proposes the future direction of gastrointestinal brain imaging and outlines a multidisciplinary approach to FGD.
BRAIN IMAGING: FUTURE DIRECTIONS
While there is some scepticism about the role of functional brain imaging for the routine assessment of FGD patients, due to its expense, poor availability, and unsuitability, its role in the systematic evaluation of the mechanisms in pain processing is not in doubt. As with all other branches of science, hypotheses should be proposed, models designed, and these models tested by experiment. This has been the approach of several somatic pain research groups and great strides in our understanding of pain physiology have resulted.17,30,42,60–64 A second important role for brain imaging will be in the evaluation of future treatments, both pharmacological and psychological.65
One of the driving forces behind research into mechanisms of gastrointestinal pain, based largely on animal studies, has been the fact that we stand on the threshold of finally having potentially effective therapeutic agents that specifically attenuate visceral hypersensitivity.66 However, the modest results so far and high placebo response rates of many drugs developed for FGD most likely reflects the heterogeneity of this group of patients.67,68 Therefore, it is imperative that we are able to differentiate the primary mechanisms of visceral hypersensitivity in FGD so that future treatment strategies can be appropriately targeted and, as discussed earlier, brain imaging alone cannot achieve this.
In order to achieve the goal of identifying subgroups of FGD patients with similar mechanisms of pain, it is important that we develop human models which explain the processes involved in the development of visceral hypersensitivity. Recently, a human model of gastrointestinal injury/inflammation induced visceral hypersensitivity has been developed. In this model, infusion of dilute hydrochloric acid into the distal oesophagus induced a prolonged reduction of pain thresholds in the proximal non-acid exposed oesophagus, and this response was exaggerated in patients with non-cardiac chest pain.69 The mechanism by which this occurs is thought to be related to sensitisation of visceral afferents within the spinal cord (central sensitisation). Furthermore, pharmacological profiling of this model has demonstrated that both prostaglandin PGE2 and N-methyl-d-aspartate receptor antagonists have a role in modulating central sensitisation.70,71 It is therefore now possible to speculate that patients with visceral hypersensitivity secondary to central sensitisation will respond to agents that block central sensitisation (anti-hyperalgesics) while those with visceral hypersensitivity secondary to hypervigilance will not respond to these drugs, although one must take into account any central effects elicited by such drugs when interpreting these data.
Using this approach, we may be able to identify subgroups of FGD patients that could then be studied with specific hypothesis testing experiments using functional brain imaging techniques. Similar profiling of patients using a combination of psychological, physiological, pharmacological, and molecular biological assessments is likely to truly enhance our understanding of pain mechanisms in FGD.
This approach is akin to that currently used by neurologists when addressing the problem of somatic central pain syndromes. Individual patients presenting in clinic undergo a battery of clinical assessments and multimodal quantative sensory testing. In addition, objective neurophysiological information regarding the transmission and processing of nociceptive information is obtained with the use of cortical evoked potentials (CEP).72 Neurophysiologists have used CEP to study somatosensory, visual, auditory, and pain pathways for over 50 years.73 This technique involves the brief presentation of a sensory stimulus which is time and phase locked to the recording of the electroencephalogram (EEG) via surface electrodes placed on the scalp. The event related signal is small in amplitude but occurs at the same moment in time following each stimulus while the large amplitude background EEG occurs randomly. In order to extract the desired signal, repeated stimuli are given and the subsequent brain activity averaged. This reduces the unwanted randomly occurring EEG while enhancing the event related CEP.74 The resultant waveform represents the brain’s response to a stimulus as it changes on a millisecond by millisecond basis.
CEP responses are reproducible and have been recorded in response to stimulation of many regions of the gastrointestinal tract (for reviews see Hobson and Aziz,56,76 and Hollerbach and colleagues75), and these responses reflect the known physiological characteristics of the afferents that innervate these regions (fig 4).77 Importantly, the gastrointestinal CEP response provides an objective correlate of perceived sensation, with the response increasing in amplitude while decreasing in latency, as the stimulation intensity increases (fig 5).78,79 This ability to record objective neurophysiological measures which correspond directly to subjective pain ratings overcomes many of the limitations of previous studies of visceral hypersensitivity.40 In addition, it has recently been shown that the CEP response to proximal oesophageal stimulation is enhanced following distal oesophageal acidification in the model of oesophageal sensitisation, indicating that CEP enables us to detect changes in the sensitivity of the central gastrointestinal pain pathway.80

Three cortical evoked potential (CEP) responses obtained from one female subject in response to electrical stimulation of the oesophagus, sigmoid colon, and rectum. It can be seen that the fact that the sigmoid is distal to the oesophagus is reflected by the increase in latency of the P1 component of the sigmoid response, due to the increased distance the afferent signal has to travel. However, it can also be noted that the rectal P1 response occurs earlier than both the oesophageal and sigmoid responses. This reflects the presence of faster conducting afferent fibres in the rectum, which is consistent with its role in maintaining continence.
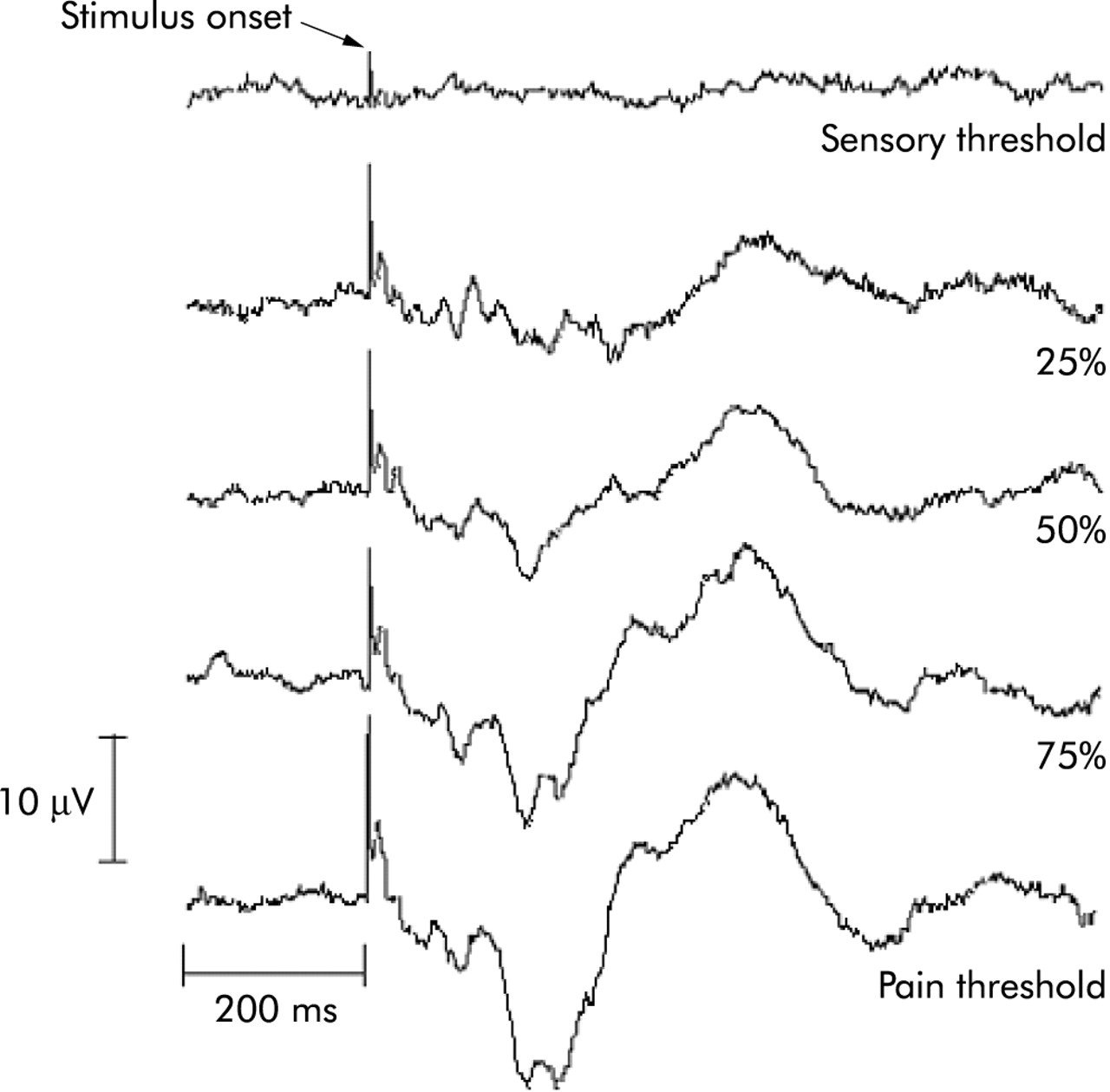
Effect of increasing rectal stimulation intensities on the subsequent cortical evoked potential (CEP) response. It can be seen that as the intensity rises from sensory threshold through to pain, there is an associated increase in the amplitude of the CEP response and a decrease in the latency of each CEP component. This provides an objective neurophysiological correlate of subjective pain reports.
CEP also has several practical advantages over other brain imaging techniques. These include the fact that the equipment needed to perform these studies is inexpensive (<£10 000), is available in most hospitals, and can be performed in individual patients as a routine clinical assessment. However, its major disadvantage is that while we know that CEP represents a summation of cortical activity related to specific stages in the cortical processing of gastrointestinal sensation and pain, the signal is distorted as it passes through the skull and scalp, making localisation of the neural correlates of the response difficult. Magnetoencephalography (MEG) is another brain imaging technique that has been used to address this issue. MEG detects the minute magnetic fields generated by groups of active cortical neurones using highly sensitive sensors known as SQUIDS (super conducting quantum interference device). Unlike the electrical signal recorded with EP, which is distorted by all of the structures that lie between the cortical source and recording electrode, the magnetic field generated by an active group of cortical neurones passes through these structures unattenuated.81 The major advantage of using MEG therefore is that it has comparable spatial resolution to PET and fMRI in addition to millisecond temporal resolution, and therefore pain related cortical activity can be imaged dynamically as it changes in real time. This is advantageous when compared with metabolic imaging techniques as neuronal activity can be recorded in specific brain regions throughout each of the temporally distinct stages of pain processing. In addition, as the signal detected with MEG is the magnetic component of the electrical field recorded with CEP, it can be used to temporally correlate activity occurring within specific cortical regions with the scalp recorded CEP.
Using this approach it has been demonstrated that the earliest oesophageal CEP component (P1), which occurs approximately 80–110 ms following oesophageal stimulation, reflects activation of cortical regions involved in processing the sensory discriminatory aspects of oesophageal pain (S1, S2, and posterior insula) while later components (N1, P2) are an amalgamation of cortical activity within regions that process both sensory discrimination and the affective/cognitive components.82,83 It is clear therefore that these later components represent secondary processing of oesophageal sensation/pain.
The relevance of these findings to FGD comes from a recent case report in which the oesophageal CEP response in two patients with non-cardiac chest pain was examined. In this case study, two male patients presented in clinic with symptoms of angina-like chest pain in the absence of any cardiac abnormality (as tested by exercise ECG and coronary angiography). Both had a previous trial of proton pump inhibitors which did not alleviate symptoms and there was no evidence of motor abnormalities or gastro-oesophageal reflux on physiological testing. Oesophageal balloon distension and electrical stimulation revealed evidence of oesophageal hypersensitivity using the method of ascending limits technique. An electrical stimulation intensity which was 75% of the difference between the subject’s sensory and pain thresholds was then used to elicit oesophageal CEP.
Figure 6 shows oesophageal CEP responses in both patients and demonstrates that these seemingly identical patients have drastically different neurophysiological profiles. Patient A demonstrates visceral hypersensitivity to oesophageal stimulation in addition to short latency large amplitude CEP responses. This is the same type of response seen in the acid model of oesophageal sensitisation and is consistent with this patient having sensitisation of the gastrointestinal afferent pathways. In contrast, patient B, while still demonstrating visceral hypersensitivity, has long latency early CEP components, indicating that the afferent signal arriving at the cortex is not exaggerated, yet the late responses, which reflect affective processing, are enhanced when compared with normal. This profile is consistent with a second proposed subgroup of non-cardiac chest pain: those that are hypervigilant of visceral sensations. In summary, this case study demonstrates that patients with non-cardiac chest pain can be subclassified on the basis of sensory responsiveness and neurophysiological profiles. While these preliminary observations hold promise, they require confirmation in a larger cohort of patients. However, if this approach is successful, then it may allow us to identify specific abnormalities within the central nervous system processing of gastrointestinal sensation in individual patients, allowing us to objectively differentiate between those with sensitised gastrointestinal afferents from those that are hypervigilant to gastrointestinal sensations. The ability to differentiate between these two groups has obvious advantages when deciding to treat them with drugs that may be anti-hyperalgesic in contrast with analgesic.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A normal cortical evoked potential (CEP) (top panel) and an oesophageal evoked potential (OEP) in two patients with non-cardiac chest pain (NCCP). Patient A demonstrated oesophageal hypersensitivity and this was associated with enhanced early and late CEP components. This profile is consistent with patient A having sensitised oesophageal afferents. Patient B demonstrated oesophageal hypersensitivity and this was associated with reduced amplitude and delayed latency of the first three CEP components. This is indicative of over reporting (that is, normal afferent transmission of a reduced stimulus intensity). However, there was a small enhancement of the late (500 ms) CEP component, which is likely to be associated with secondary pain processing. This patient does not have sensitised oesophageal afferents and is hypervigilant of oesophageal stimuli.
While these data come from studies of the oesophagus, CEP can be recorded from the duodenum, rectum, and sigmoid,77 and therefore these findings are likely to be replicable in other FGDs such as functional dyspepsia and IBS. The importance of these findings is to underline that FGD comprises a heterogeneous group of patients, and only when we have defined the phenotype of these conditions and identified groups of patients with specific CNS abnormalities will it be possible to perform clinical brain imaging studies in homogenous FGD populations, aimed at answering specific hypotheses.
CONCLUDING REMARKS
In conclusion, we believe that while functional brain imaging has considerable potential in helping us to understand gastrointestinal pain neurophysiology and in the development of models in healthy subjects that explore the modulatory effects of psychological influences and injury/inflammation induced gastrointestinal hypersensitivity, their application to the study of FGD is somewhat premature. Furthermore, the availability of functional brain imaging techniques provides an unparalleled opportunity to make fundamental advances in our understanding of the mechanisms of pain in FGD. However, this will only occur if we use brain imaging as a complimentary investigative tool in conjunction with psychological, physiological, pharmacological, and biological profiling of FGD patients.