Article Text

Abstract
Background and aims: The exact nature of the epithelial barrier defect in Crohn’s disease remains to be elucidated. Previously we showed increased permeability to proteins in ileal Crohn’s disease. Our aims were to study if this barrier defect (a) involves endocytotic uptake of antigens and (b) is related to low grade inflammation not detectable by histology.
Methods: Macroscopically normal segments of distal ileum of Crohn’s disease patients (n = 10) were subgrouped into non-inflamed (histologically unaffected) and slightly inflamed tissues and studied in Ussing chambers, with normal ileal specimens from colon cancer patients (n = 9) as controls. Endocytotic uptake into enterocytes of the protein antigen horseradish peroxidase was assessed by measuring the area of horseradish peroxidase containing endosomes in electron photomicrographs. Mucosal tumour necrosis factor α (TNF-α) mRNA was quantified using real time polymerase chain reaction. For comparison, the effects of low doses of TNF-α on endosomal uptake of horseradish peroxidase were studied in cultured T84 cells grown on filter supports.
Results: The area of horseradish peroxidase containing endosomes was increased (p<0.001) in enterocytes of non-inflamed ileum of Crohn’s disease (2.8 (0.7) μm2/300 μm2) compared with control ileum (0.6 (0.06)). In non-inflamed mucosa, a significant association between endosomal uptake and mucosal expression of TNF-α mRNA (p = 0.03) was found. Low concentrations of TNF-α (0.25–1.0 ng/ml) enhanced the endosomal uptake of horseradish peroxidase in polarised T84 cells, without affecting transepithelial electrical resistance.
Conclusions: Our findings suggest increased endosomal uptake of antigens in ileal Crohn’s disease that may be mediated by TNF-α. These data highlight the transcellular route of antigen uptake in barrier dysfunction and implicate the interaction between epithelial cells and the innate immune system in the development of mucosal inflammation.
- CD, Crohn’s disease
- Isc, short circuit current
- HRP, horseradish peroxidase
- RT-PCR, reverse transcripion-polymerase chain reaction
- PD, potential difference
- TER, transepithelial electrical resistance
- TNF-α, tumour necrosis factor α
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- electron microscopy
- Crohn’s disease
- endocytosis
- tumour necrosis factor α
- permeability
- T84 cell line
Statistics from Altmetric.com
- CD, Crohn’s disease
- Isc, short circuit current
- HRP, horseradish peroxidase
- RT-PCR, reverse transcripion-polymerase chain reaction
- PD, potential difference
- TER, transepithelial electrical resistance
- TNF-α, tumour necrosis factor α
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
The last decade of research has given us insights into the critical role of the bacterial flora and the importance of dysregulation of the mucosal immune system in intestinal inflammation. The intestinal epithelial barrier is the interface between these two environments, and sufficient disruption of the barrier can initiate and perpetuate inflammation.1–3
Increased intestinal permeability to medium sized molecules, reflecting paracellular permeability, is a characteristic of intestinal inflammation. In patients with Crohn’s disease (CD), increased gut permeability can predict relapse and may precede histological inflammation.4,5 Several studies have demonstrated a subgroup of first degree relatives with a disrupted intestinal barrier,6,7,8,9,10 and a case report has shown development of CD in a relative eight years after a finding of increased intestinal permeability.11 Therefore, increased intestinal permeability has been suggested as a factor of pathogenic importance in CD. On the other hand, increased permeability in CD is related to disease activity,4,5 and recently Suenaert and colleagues12 reported that treatment with anti-tumour necrosis factor α (TNF-α) antibody largely restores the gut barrier in CD. This could imply that increased permeability in CD is secondary to the inflammatory process. Moreover, Yacyshyn and colleagues13 found coexistence of increased permeability and increased expression of activation markers on peripheral blood lymphocytes in CD relatives, suggesting that the disturbed permeability in relatives of CD is also associated with an activated immune system. The recent findings that 50% of healthy CD relatives show signs of subclinical intestinal inflammation14 also support this concept. This underlines the need for studies of immune regulation of the mucosal barrier in the pathophysiology of CD.
Data from animal models of intestinal inflammation have revealed that conditions with an increased intestinal permeability to medium sized molecules in vivo show a combination of increased tight junction permeability and enhanced flux of proteins (mainly transcellular uptake) when studied in Ussing chambers in vitro.15,16 A combination of enhanced permeability to proteins and tight junction dysfunction was also found in ileal CD mucosa studied in vitro,17,18 as well as in ulcerative colitis.19,20 While regulation of paracellular permeability has received much attention in recent years,21–25 little is known about transcellular uptake of antigens in human inflammatory bowel disease.
TNF-α is one of the most important proinflammatory mediators in CD, and treatment with anti-TNF-α has become a cornerstone in CD treatment. Moreover, an important role of TNF-α in regulating intestinal barrier function (transcellular and paracellular route) has been demonstrated by coculture of intestinal cell lines with human mononuclear cells from patients with active CD,26 as well as from patients with human immunodeficiency virus enteropathy,27 cow’s milk allergy,28 and monocytes activated by bacterial superantigen.29 In a previous study,18 we showed increased permeability to ovalbumin in non-inflamed ileum of CD, possibly via the transcellular route. From this study two important questions have emerged: Does the barrier defect in ileal CD involve endocytotic uptake of antigens as a first step of transcytosis? And, if so, is this related to low grade inflammation (not detectable by histology) and TNF-α production?
The present study shows that increased endosomal uptake of the protein antigen horseradish peroxidase (HRP) into enterocytes was correlated with expression of TNF-α mRNA in histologically unaffected ileal mucosa of CD. Moreover, low concentrations of TNF-α induced increased epithelial endocytosis of HRP in polarised T84 cells. Our findings suggest that mucosal barrier dysfunction in the pathogenesis of CD includes the transcellular route and may be secondary to release of TNF-α.
METHODS
Experiments in human tissue
Patients and ethics
The study comprised 10 patients operated on for CD (table 1), with nine patients undergoing surgery for colonic cancer as controls. CD patients (seven women, three men), had a median age of 32 (range 21–59) years and underwent elective surgery for ileal (n = 8) or ileocolonic (n = 2) CD at McMaster University Medical Centre. Disease duration prior to the present surgery was 4 (range 1–15) years; eight were primary resections and two re-resections. Six patients were on maintenance treatment with azathioprine or 6-mercaptopurine and four were receiving corticosteroids with or without mesalasine. Indications for surgery were obstructive symptoms in five cases, previous intra-abdominal abscesses in three cases, and enterocolonic fistulas in two patients. Ileocaecal resection was performed in eight patients whereas combined ileal and more extended colonic resection was done in the remaining two. Nine patients (six men) operated on for right hemicolectomy (n = 5) or subtotal colectomy (n = 4) for colon cancer, aged 55 (range 45–81) years, served as controls. The colon cancer patients had no evidence of generalised disease. None had received preoperative chemotherapy or radiotherapy.
Characteristics of the patients and histological assessment of ileal specimens in the 10 patients with Crohn’s disease included in the study
The study was approved by the Ethics Committee, Faculty of Health Sciences, McMaster University, and was conducted according to the Declaration of Helsinki.
Ussing chamber experiments
Surgical specimens of distal ileum from patients with CD (n = 10) were studied, with normal ileal mucosal specimens from colon cancer patients (n = 9) as controls. Immediately after removal of the vascular supply to the distal ileum, segments were put into cold oxygenated Krebs’ buffer and then macroscopically reviewed by a pathologist (CS). Macroscopically non-involved segments were stripped of muscle layers and 2–4 adjacent pieces from each patient were mounted in modified Ussing chambers (W-P Instruments, Narco Scientific, Mississauga, Ontario, Canada). The chamber opening exposed 0.6 cm2 of tissue surface area to 8 ml of circulating oxygenated Krebs buffer at 37°C. The buffer contained (in mM) 115 NaCl, 1.25 CaCl2, 1.2 MgCl2, 2.0 KH2PO4, and 25 NaHCO3, pH 7.35±0.02. In addition, the serosal buffer contained 10 mM glucose as an energy source, osmotically balanced by 10 mM mannitol in the mucosal buffer. Agar salt bridges were used to monitor the potential difference (PD) across the tissue and to inject the required short circuit current (Isc) to maintain a zero potential difference, as registered via an automated voltage clamp (W-P Instruments). Isc (μA/cm2) was recorded continuously by a computer and tissue conductance was calculated according to Ohm’s law and expressed as mS/cm2. Baseline values for Isc (active ion secretion) and conductance (passive ion permeability) were assessed at equilibrium, 15 minutes after mounting the tissues. At this time point, HRP was added to the luminal chamber. In the five minute experiments, tissues were then fixed in glutaraldehyde without further electrical measurements. In the 20 minute experiments, Isc was studied at baseline, following transmural nerve stimulation, and after addition of glucose (10 mM) to the luminal side. A PD of 6 mV with glucose in the luminal chamber was previously determined as a parameter of viability.18,30 Therefore, a prerequisite for inclusion in the present study was a rise in PD to no less than 6 mV after luminal addition of glucose in the 20 minute experiments.
Epithelial uptake of HRP
Endocytotic uptake into epithelial cells was assessed in ileal segments after five minutes and 20 minutes of luminal exposure to HRP. At these time points, tissues were removed from the chambers and immediately fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4) for two hours at 22°C, rinsed for 18 hours (4°C) with 0.05 M Tris buffer (pH 7.6), and washed three times, five minutes each time. Methods for HRP product identification were modified from Graham and Karnovsky.31,32 In brief, peroxidase activity was demonstrated by incubating tissues for 15 minutes in 0.5 mg/ml diaminobenzidine in 0.05 M Tris buffer (pH 7.6, 22°C), followed by 15 minutes of incubation in the same buffer containing 0.01% H2O2. Subsequently, tissues were processed for transmission electron microscopy. Quantitative analysis of HRP uptake in intracellular endosomes and paracellular HRP transport were performed on coded high magnification photomicrographs, 12 per patient. At the five minute time point, photomicrographs from six control patients, six non-inflamed CD, and four inflamed CD were assessed. At 20 minutes, photomicrographs from three controls and five CD patients (two non-inflamed, three slightly inflamed) were studied. The total area of HRP containing endosomes within enterocytes from the mid-villus region was determined in an area of 300 μm2 in the apical region of the cells, using a computerised image analysis system (Kontron Mop Videoplan, Kontron, Eching, Germany).33,34 Preliminary studies determined that endogenous peroxidase was not evident in gut epithelial cells of either controls or CD patients.
Quantification of tissue TNF-α mRNA using real time PCR
Samples for polymerase chain reaction (PCR) were snap frozen and stored at −70°C until subsequent analyses. For RNA extraction, the tissue was pulverised in liquid N2 and RNA was extracted using Trizol (Invitrogen, Carlsbad, California, USA) according to the product protocol. RNA concentration was determined by spectrophotometry at OD 260 nm and the quality of RNA by the presence of intact 18S and 28S rRNA after electrophoresis on an ethidium bromide stained 1% agarose gel. After RNA isolation, 10 μg of each sample were treated with DNase I to remove genomic DNA using the DNA free kit (Ambion, Austin, Texas, USA). Purified samples were then re-quantified and 1 μg utilised for reverse transcription with the RETROscript kit (Ambion). After the completed reaction each sample was diluted to 200 μl with nuclease free water (Ambion) and stored at −20°C until use. For real time PCR, each well contained human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) forward and reverse primers (10 μm, 1.25 μl/reaction; Applied Biosystems, Foster City, California, USA), GAPDH probe (VIC reporter, 20 μm, 1.25 μl/reaction), Taqman Universal PCR master mix (12.5 μl; Applied Biosystems), TNF-α primers and probe (human TNF-α Pre-developed Assay Reagent, FAM reporter, catalogue No 4327055; Applied Biosystems,) 5 μl of nuclease free water, and 5 μl of sample cDNA. For each run, a non-transcribed sample of RNA reaction mix as well as a reverse transcription (RT) reaction mix with no cDNA present was utilised as a control. All samples were run in triplicate. PCR was done in the ABI PRISM 6700 Sequence Detection system. Gene expression was quantified relative to expression of GAPDH. TNF-α mRNA expression in control patients was averaged to obtain baseline expression, and expression in CD tissues was then divided by control expression and the results presented as percentage of control.
Histology
Tissue was taken adjacent to that studied in the Ussing chamber, fixed in 4% formaldehyde, embedded in paraffin, sectioned, and stained with haematoxylin-eosin. Cases were reviewed by a pathologist (CS) who was blinded with respect to the type of experiment and underlying diagnosis. Each case was assessed for degree of polymorphonuclear leucocyte infiltration in the lamina propria and epithelium, respectively, lymphocyte infiltration (chronic inflammation), mucosal atrophy, and mucosal oedema. This was performed utilising semiquantitative scales: 0, normal appearance; 1, mild changes; 2, moderate changes; and 3, severe changes. A histology score (0–15) was created from the sum of these variables in each specimen (table 1). Control patients had normal appearing ileal mucosa, with histology scores of 0–1. CD cases within the non-inflamed group had a normal macroscopic and microscopic appearance, and had scores of 0–2 when scrutinised. In the slightly inflamed group, histology score was 3–6 but all specimens had an intact mucosa (that is, no ulcers or erosions were observed).
Experiments using the T84 cell line
Cell culture
T84 epithelial cells were seeded (106 cells) on tissue culture treated Transwell filter supports (0.4 µm pore size, 1.2 cm2 surface area; Costar, Cambridge, Massachusetts, USA). Culture medium consisted of a 1:1 mixture of DMEM and Ham’s F-12 nutrient mixture (Gibco, Grand Island, New York, USA) supplemented with 10% (vol/vol) heat inactivated fetal bovine serum (Cansera International, Rexdale, Ontario, Canada), 1.5% HEPES (Gibco), and 2% penicillin-streptomycin (Gibco). Cells were grown at 37°C in 5% CO2 for seven days to attain polarised epithelial monolayers with a transepithelial resistance (TER) of >1000 Ωcm2. Monolayer resistance was monitored throughout the experimental period using a Millicell-ERS system (Millipore, Bedford, Massachusetts, USA). TER serves as a measure of the ability of T84 cells to restrict passive movement of ions.
Treatment of monolayers with TNF-α
Confluent monolayers were cultured for 48 hours before the flux experiments with 0–10.0 ng/ml of recombinant human TNF-α (R&D Systems, Minneapolis, Minnesota, USA) added to the basal compartment (volume 1.5 ml) of the Transwell unit. TNF-α doses were chosen based on previous studies showing only minor effects on TER in these concentrations.35–37
Transepithelial transport of HRP
HRP (molecular weight 44 000, type VI-A; Sigma, St Louis, Missouri, USA) was added to the apical side of the monolayers at a concentration of 10−5 M, and monolayers were incubated for two hours at 37°C. Media samples were obtained from the basal compartments and assayed for HRP as described above.
Electron microscopy
To determine the route and magnitude of transepithelial transport of HRP, T84 monolayers exposed to HRP were processed for transmission electron microscopy. Briefly, 120 minutes after HRP addition, T84 monolayers were fixed with 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4) and further processed as described above. The 120 minute time period was chosen to enable simultaneous measurement of HRP containing endosomes, HPR fluxes, and the presence or absence of HRP in the tight junctions. An observer unaware of the treatment groups (P-CY) analysed random photomicrographs. For each experimental group, intracellular HRP was quantified by determining the total endosomal HRP area in 300 μm2 windows (12 windows/group, three separate experiments). Tight junctions and paracellular regions were also reviewed in a blinded fashion for the presence of HRP.
Statistics
Results are expressed as means (SEM) unless otherwise stated. Analyses were performed with ANOVA, the Student’s t test, or simple linear regression, as applicable. Differences with p<0.05 were considered significant.
RESULTS
CD mucosa shows low baseline Isc but equal viability as control tissue
Ileal mucosa from CD (n = 32 specimens from 10 patients) had lower baseline Isc (54 (6) μA/cm2) compared with control ileum (103 (7) μA/cm2, n = 23 specimens from nine patients; p<0.01). There were however no differences between the patient groups with respect to the rise in Isc following transmural electrical stimulation, reflecting neurally mediated activation of anion secretion (CD v control: 41 (9) v 39 (7) μA/cm2; p>0.05),38 or in the Isc response to luminal glucose, activating glucose coupled sodium absorption (CD v control: 85 (20) v 101 (27) μA/cm2; p>0.05). Initial baseline conductance was also similar in the two study groups (CD v control: 24.4 (1.9) v 27.3 (1.7) mS/cm2; p>0.05). These findings indicate similar integrity and viability of ileal tissues obtained from patients with CD and colon cancer controls.
Endosomal uptake of HRP is increased in CD
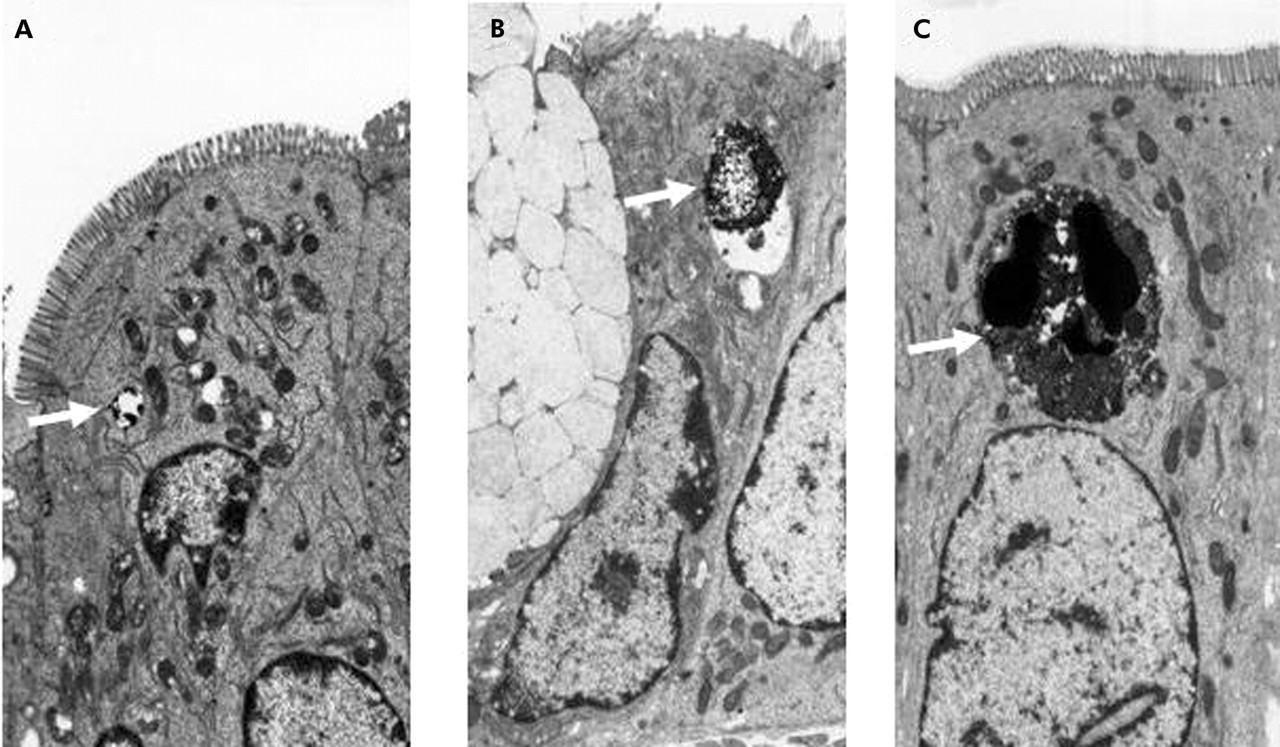
Endosomal uptake studied after five minutes of luminal exposure showed HRP in small endosomes in the ileum from control patients (fig 1A) whereas in CD, larger HRP containing endosomes were found (fig 1B, C). When quantified by image analysis, HRP endosome area was substantially higher in non-inflamed ileum of CD (2.8 (0.7) μm2/300 μm2 apical cell area, n = 6 patients) than in control ileum (0.6 (0.06) μm2/300 μm2, n = 6 patients, p<0.001) (fig 2A), and was even further increased in the slightly inflamed CD specimens (4.4 (0.4) μm2/300 μm2, n = 4 patients) (fig 2A). Linear regression showed a significant association between histology score and area of HRP containing endosomes (p<0.01; fig 2B). Endosomal area was increased to a similar extent after 20 minutes of luminal exposure to HRP (control ileum (n = 3 patients) 0.7 (0.2); CD ileum (n = 5 patients) 2.2 (0.5) μm2/300 μm2; p<0.05). No HRP was seen in paracellular spaces.

Increased endosomal uptake of horseradish peroxidase (HRP) in Crohn’s disease (CD). Representative transmission electron photomicrographs of mid-villus ileal epithelium from a control patient (A) and the non-inflamed ileum of patients with CD (B, C), fixed five minutes after addition of HRP to the luminal buffer in Ussing chambers. HRP containing endosomes are indicated by arrows. Note the difference in endosome area between CD and control patients (see fig 2). Original magnification ×5000.

Increased endosomal uptake of horseradish peroxidase (HRP) in Crohn’s disease (CD). Quantification of endosomal uptake of HRP in ileal enterocytes of control patients (n = 6), non-inflamed CD (n = 6), and slightly inflamed mucosa of CD patients (n = 4). (A) Data show total area of HRP containing endosomes within enterocytes determined in an area of 300 μm2 in the apical region of the cells; 12 randomly chosen cells assessed per patient. Values are means (SEM). **Increased compared with controls, p<0.01. (B) Linear regression using the same observations as in (A), with histology score (see table 1) as the independent variable and endosomal uptake as the dependent variable: r = 0.72, t value 3.88, p<0.01.
TNF-α mRNA is increased in CD mucosa and is associated with endosomal uptake
As TNF-α is an important proinflammatory cytokine in CD and a regulator of mucosal barrier function, we assessed tissue levels of TNF-α by real time RT-PCR. TNF-α mRNA levels in ileal tissue of CD patients were increased to 323% (207–658%) (median, interquartile range) of control ileum (p<0.001; fig 3A). In the subgroup of patients with non-inflamed mucosa (histology score 0–2), endosomal uptake of HRP at five minutes increased with tissue expression of mRNA for TNF-α (p = 0.03; fig 3B).

(A) Relative expression of tumour necrosis factor α (TNF-α) mRNA in ileal tissue of Crohn’s disease (CD) and control patients, as determined by real time reverse transcription-polymerase chain reaction. Horizontal line denotes median value. (B) Linear regression using TNF-α expression in controls and non-inflamed CD as the independent variable and endosomal uptake as the dependent variable: r = 0.71, t value 2.64, p<0.03.
TNF-α increases HRP flux and endosomal uptake in T84 cell lines
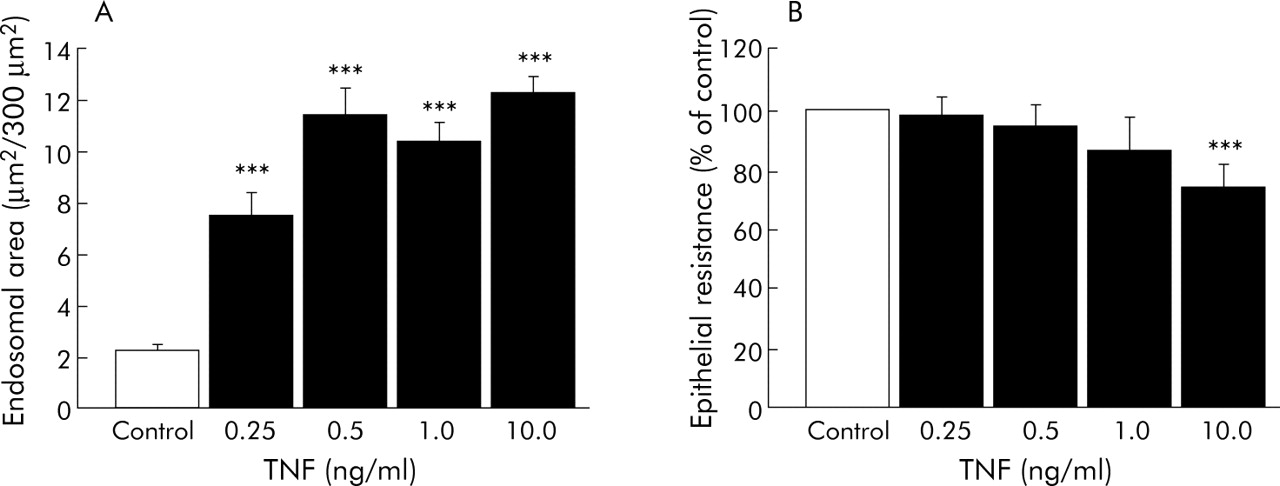
To elucidate whether the association between mucosal TNF-α and endosomal uptake is of mechanistic importance, the effects of low concentrations of TNF-α (0.25–10.0 ng/ml) were studied in polarised T84 cells (n = 9 filters in each group, three separate experiments). TNF-α caused a dose dependent increase in the area of HRP containing endosomes seen by transmission electron microscopy (fig 4A). Moreover, HRP flux over two hours increased dose dependently (TNF-α 1.0 ng/ml: 211 (50)%; TNF-α 10.0 ng/ml: 304 (20)% of control, p<0.05). On the other hand, TER at 48 hours was unaffected by TNF-α 0.25–1.0 ng/ml, and TNF-α 10.0 ng/ml caused only a moderate decrease in TER (fig 4B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dose-response effects of tumour necrosis factor α (TNF-α) in T84 cell lines. (A) Area of horseradish peroxidase (HRP) containing endosomes in epithelial cells. Cells were grown on filter supports with TNF-α (0.25–10.0 ng/ml) for 48 hours and exposed to HRP in the luminal compartment for two hours. Data show total area of HRP containing endosomes within enterocytes determined in an area of 300 μm2 in the apical region of the cells; 12 randomly chosen cells assessed per group per experiment. (B) Transepithelial electrical resistance in T84 cells grown with TNF-α (0.25–10.0 ng/ml) for 48 hours. Data are from three separate experiments with 3–4 filters studied in each group on each occasion; analysis done with n = 3, ANOVA with Fisher’s post hoc test. ***p<0.001 compared with control experiments.
DISCUSSION
The results of the present study suggest a TNF-α-dependent increase in endosomal uptake of protein antigens into enterocytes of histologically non-involved ileal mucosa of CD patients. Moreover, in the human colonic cell line, T84, low doses of TNF-α affected endosomal uptake of proteins rather than paracellular permeability. This may imply that enhanced endocytosis precedes the previously recognised increase in paracellular permeability in CD but is none the less secondary to immune activation. Our findings explain previous studies of increased permeability to protein antigens in ileal CD,18,19,39 and also highlight the importance of studying the interaction between the mucosal immune system and epithelial function in IBD pathogenesis.
The pivotal role of TNF-α in CD pathogenesis has become apparent in recent years with the sometimes dramatic effects of anti-TNF-α treatment.40–42 TNF-α is produced in excessive amounts by macrophages and mast cells in CD mucosa,43–45 and a role for TNF-α released from immune cells has been implicated in the regulation of epithelial barrier function in CD,26 as well in other settings of intestinal inflammation.27,28 Moreover, the recent report of Suenaert and colleagues12 suggested TNF-α in the regulation of in vivo intestinal permeability in patients with CD. In the present study, we found a significant association between TNF-α mRNA levels and increasing endocytosis of HRP into enterocytes of non-inflamed ileal mucosa. In addition, experiments using T84 cells, with TNF-α concentrations in physiological doses (that is, in the lower limits of what has previously been used in cell lines and of the same order of magnitude as spontaneous secretion from human intestinal tissue46) supported TNF-α as a potential regulator of endosomal uptake of antigens in enterocytes. These novel findings further emphasise the role of TNF-α in the signalling pathways between antigens, epithelial cells, and immune cells in the modulation of gut physiological and pathophysiological reactions in inflammatory bowel disease.
We previously showed increased permeability to ovalbumin in the non-involved ileum of patients with CD, and indirect evidence suggested transcytosis of proteins.18 To test this hypothesis, the present study focused on detection of endosomal uptake into ileal enterocytes. We were able to verify increased uptake of antigenic proteins into the epithelial cells of non-involved ileum, a previously unproven defect in epithelial barrier function in CD. A possible interpretation is that enhanced endosomal uptake, once triggered by TNF-α signalling from the lamina propria, could result in increased transcytosis of intact proteins or increased presentation of antigen fragments by the MHC complexes of enterocytes to immunocytes of the lamina propria.47 These events could then serve as accelerators of the process in the early stages of mucosal inflammation. The additional increase in endocytosis in inflamed mucosa, which was unrelated to mRNA levels of TNF-α, implies that in more overt inflammation other inflammatory mediators are important in regulating endocytotic uptake into enterocytes (for example, neutrophil released oxygen and nitrogen radicals).48 The immune response to an endocytosed antigen is regulated by targeting to late endosomes containing lysosome associated membrane protein 1 and MHC class II.49 Further studies are needed to determine regulation of the intracellular trafficking of endosomal proteins within enterocytes of ileal CD.
An alternative explanation for our findings could be that TNF-α induced an increase in apoptosis of epithelial cells,50 with increased epithelial turnover and more immature cells (with more active endocytosis) reaching the absorptive part of the villi. This could result in increased paracellular leakage at apoptotic foci, as has been described in ulcerative colitis.20 These leaking foci would not be detected with the conventional resistance measurements used in the present study.51 Further studies are needed to address this point.
In recent years it has become evident that the epithelium is in constant interplay with luminal bacteria and dietary antigens, as well as with the mucosal immune system.52 One important contributing factor is the genetic susceptibility of CD, as recently highlighted by the discovery of mutations in the NOD2/CARD15 gene in 30% of CD patients.53,54 This could lead to reduced protection against luminal bacteria55 and a hyperreactive innate immune system, giving an elevated background level of TNF-α with epithelial barrier dysfunction as a consequence.26
The control group of colon cancer patients employed in this study was substantially older than the CD patients but intestinal permeability does not seem to be affected by aging.4,56 An increase in intestinal permeability has been found in advanced malignancy or related to chemotherapy.4 However, the cancer patients included in the present study had no signs of generalised disease, did not receive chemotherapy, and were in a good nutritional condition. Most CD patients were receiving treatment with corticosteroids and mesalazine. In experimental intestinal inflammation these drugs seem to tighten the barrier.57,58 In the present study, there were no differences in HRP uptake between patients with and without corticosteroid treatment. Therefore, medication is not likely to explain the differences in endosomal antigen uptake observed between the patient groups.
Samples taken close to the specimens studied in the Ussing chambers, as well as the resection margins from all patients, were scrutinised by histopathology. The “non-inflamed” CD group had a normal macroscopic and light microscopy appearance when reviewed for clinical purposes but had scores of 0–2 (and increased TNF-α expression) when scrutinised in more detail. This indicates inflammatory load in clinically non-involved areas, and supports the concept of barrier dysfunction being secondary to low grade inflammation. Mucosal specimens from CD patients also demonstrated a low baseline Isc, probably reflecting reduced net ion pump activity. It was recently shown that TNF-α reduces epithelial cell secretory function by inhibiting Na+/K+-ATPase activity by a specific cytokine effect not implying reduced viability.59 Our findings regarding electrophysiology should however be viewed with caution as it was previously shown that conventional methods for measurements of Isc and TER in inflamed human tissue are handicapped by an increase in subepithelial resistance which overestimates epithelial resistance and underestimates Isc.51 Conventional methods are nevertheless adequate in reflecting changes in Isc, and in the present study, we found similar epithelial secretory function in the patient groups, as assessed by measuring Isc changes to transmural electric nerve stimulation, and absorptive function, studied by Isc response to glucose activation of Na+/glucose co-transporters, in mucosa from CD and control patients.
In summary, we have shown for the first time that endosomal uptake of protein antigens is increased in histologically non-involved ileum of CD, and may be regulated by TNF-α. These novel findings lend further support to the importance of the transcellular route of antigen uptake in the barrier dysfunction of CD, and underline the importance of immune-epithelial interaction in the development of mucosal inflammation. In addition, these findings may give additional explanations for the positive effects of anti-TNF-α treatment in patients suffering from CD.
Acknowledgments
Funded by a Group Grant from the Crohn’s and Colitis Foundation of Canada (MP, DM, PS, JS), the Broad Medical Research Program of the Eli and Edythe L Broad Foundation (JS), and the Swedish Research Council (JS). At the time of the human tissue experiments, JS was a postdoctoral fellow sponsored by the Canadian Institutes of Health Research. PS is the recipient of a Canadian Research Chair in Gastrointestinal Disease.